19. Adaptive Specific Host Defenses
19.3 T Lymphocytes and Cellular Immunity
Learning Objectives
- Describe the process of T-cell maturation and thymic selection
- Explain the genetic events that lead to diversity of T-cell receptors
- Compare and contrast the various classes and subtypes of T cells in terms of activation and function
- Explain the mechanism by which superantigens effect unregulated T-cell activation
As explained in Overview of Specific Adaptive Immunity, the antibodies involved in humoral immunity often bind pathogens and toxins before they can attach to and invade host cells. Thus, humoral immunity is primarily concerned with fighting pathogens in extracellular spaces. However, pathogens that have already gained entry to host cells are largely protected from the humoral antibody-mediated defences. Cellular immunity, on the other hand, targets and eliminates intracellular pathogens through the actions of T lymphocytes, or T cells (Figure 19.13). T cells also play a more central role in orchestrating the overall adaptive immune response (humoral as well as cellular) along with the cellular defences of innate immunity.

T Cell Production and Maturation
T cells, like all other white blood cells involved in innate and adaptive immunity, are formed from multipotent hematopoietic stem cells (HSCs) in the bone marrow (see Figure 18.13). However, unlike the white blood cells of innate immunity, eventual T cells differentiate first into lymphoid stem cells that then become small, immature lymphocytes, sometimes called lymphoblasts. The first steps of differentiation occur in the red marrow of bones (Figure 19.14), after which immature T lymphocytes enter the bloodstream and travel to the thymus for the final steps of maturation (Figure 19.15). Once in the thymus, the immature T lymphocytes are referred to as thymocytes.
The maturation of thymocytes within the thymus can be divided into tree critical steps of positive and negative selection, collectively referred to as thymic selection. The first step of thymic selection occurs in the cortex of the thymus and involves the development of a functional T-cell receptor (TCR) that is required for activation by APCs. Thymocytes with defective TCRs are removed by negative selection through the induction of apoptosis (programmed controlled cell death). The second step of thymic selection also occurs in the cortex and involves the positive selection of thymocytes that will interact appropriately with MHC molecules. Thymocytes that can interact appropriately with MHC molecules receive a positive stimulation that moves them further through the process of maturation, whereas thymocytes that do not interact appropriately are not stimulated and are eliminated by apoptosis. The third and final step of thymic selection occurs in both the cortex and medulla and involves negative selection to remove self-reacting thymocytes, those that react to self-antigens, by apoptosis. This final step is sometimes referred to as central tolerance because it prevents self-reacting T cells from reaching the bloodstream and potentially causing autoimmune disease, which occurs when the immune system attacks healthy “self” cells.
Despite central tolerance, some self-reactive T cells generally escape the thymus and enter the peripheral bloodstream. Therefore, a second line of defence called peripheral tolerance is needed to protect against autoimmune disease. Peripheral tolerance involves mechanisms of anergy and inhibition of self-reactive T cells by regulatory T cells. Anergy refers to a state of non-responsiveness to antigen stimulation. In the case of self-reactive T cells that escape the thymus, lack of an essential co-stimulatory signal required for activation causes anergy and prevents autoimmune activation. Regulatory T cells participate in peripheral tolerance by inhibiting the activation and function of self-reactive T cells and by secreting anti-inflammatory cytokines.
It is not completely understood what events specifically direct maturation of thymocytes into regulatory T cells. Current theories suggest the critical events may occur during the third step of thymic selection, when most self-reactive T cells are eliminated. Regulatory T cells may receive a unique signal that is below the threshold required to target them for negative selection and apoptosis. Consequently, these cells continue to mature and then exit the thymus, armed to inhibit the activation of self-reactive T cells.
It has been estimated that the three steps of thymic selection eliminate 98% of thymocytes. The remaining 2% that exit the thymus migrate through the bloodstream and lymphatic system to sites of secondary lymphoid organs/tissues, such as the lymph nodes, spleen, and tonsils (Figure 19.15), where they await activation through the presentation of specific antigens by APCs. Until they are activated, they are known as mature naïve T cells.
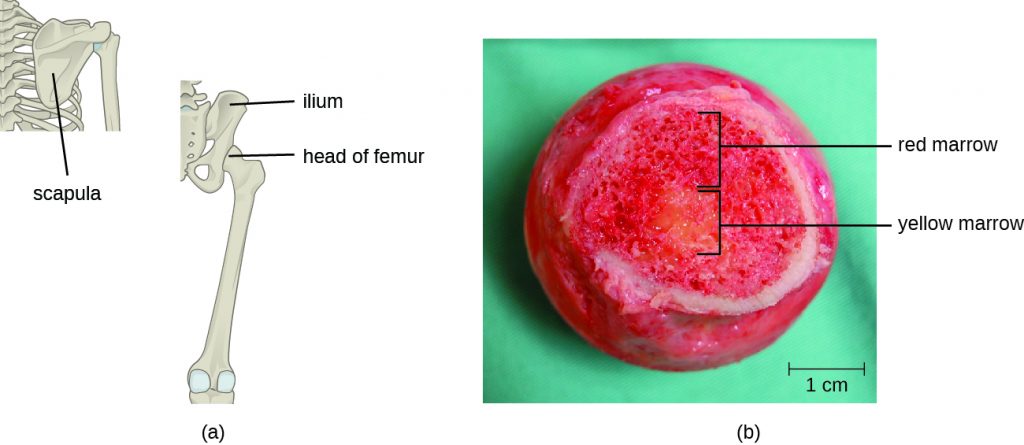

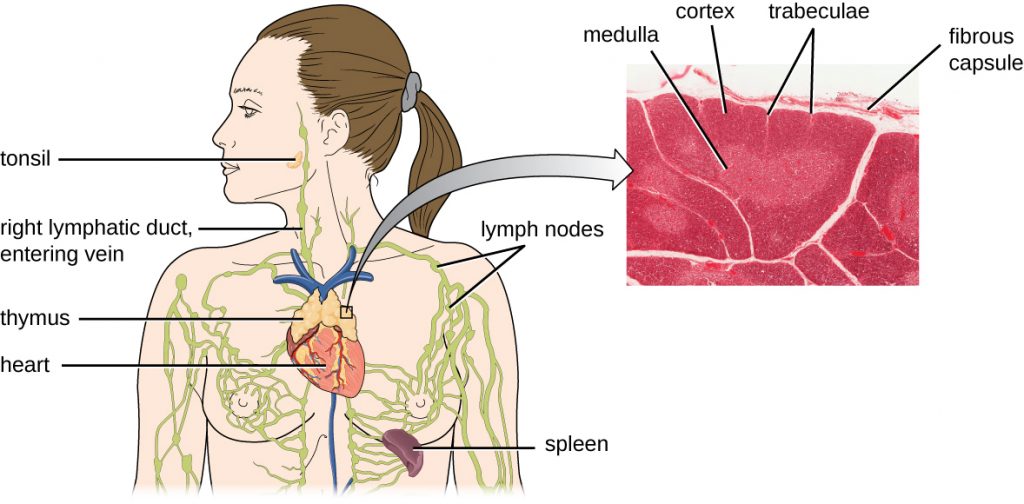
What anatomical sites are involved in T cell production and maturation?
What are the three steps involved in thymic selection?
Why are central tolerance and peripheral tolerance important? What do they prevent?
Classes of T Cells
T cells can be categorized into three distinct classes: helper T cells, regulatory T cells, and cytotoxic T cells. These classes are differentiated based on their expression of certain surface molecules, their mode of activation, and their functional roles in adaptive immunity (Table 19.1).
All T cells produce cluster of differentiation (CD) molecules, cell surface glycoproteins that can be used to identify and distinguish between the various types of white blood cells. Although T cells can produce a variety of CD molecules, CD4 and CD8 are the two most important used for differentiation of the classes. Helper T cells and regulatory T cells are characterized by the expression of CD4 on their surface, whereas cytotoxic T cells are characterized by the expression of CD8.
Classes of T cells can also be distinguished by the specific MHC molecules and APCs with which they interact for activation. Helper T cells and regulatory T cells can only be activated by APCs presenting antigens associated with MHC II. In contrast, cytotoxic T cells recognize antigens presented in association with MHC I, either by APCs or by nucleated cells infected with an intracellular pathogen.
The different classes of T cells also play different functional roles in the immune system. Helper T cells serve as the central orchestrators that help activate and direct functions of humoral and cellular immunity. In addition, helper T cells enhance the pathogen-killing functions of macrophages and NK cells of innate immunity. In contrast, the primary role of regulatory T cells is to prevent undesirable and potentially damaging immune responses. Their role in peripheral tolerance, for example, protects against autoimmune disorders, as discussed earlier. Finally, cytotoxic T cells are the primary effector cells for cellular immunity. They recognize and target cells that have been infected by intracellular pathogens, destroying infected cells along with the pathogens inside.
Table 19.1. Classes of T Cells and Their Properties
| Classes of T Cells | |||
|---|---|---|---|
| Class | Surface CD Molecules | Activation | Functions |
| Helper T cells | CD4 | APCs presenting antigens associated with MHC II | Orchestrate humoral and cellular immunity |
| Involved in the activation of macrophages and NK cells | |||
| Regulatory T cells | CD4 | APCs presenting antigens associated with MHC II | Involved in peripheral tolerance and prevention of autoimmune responses |
| Cytotoxic T cells | CD8 | APCs or infected nucleated cells presenting antigens associated with MHC I | Destroy cells infected with intracellular pathogens |
- What are the unique functions of the three classes of T cells?
- Which T cells can be activated by antigens presented by cells other than APCs?
T-Cell Receptors
For both helper T cells and cytotoxic T cells, activation is a complex process that requires the interactions of multiple molecules and exposure to cytokines. The T-cell receptor (TCR) is involved in the first step of pathogen epitope recognition during the activation process.
The TCR comes from the same receptor family as the antibodies IgD and IgM, the antigen receptors on the B cell membrane surface, and thus shares common structural elements. Similar to antibodies, the TCR has a variable region and a constant region, and the variable region provides the antigen-binding site (Figure 19.16). However, the structure of TCR is smaller and less complex than the immunoglobulin molecules (Figure 19.5). Whereas immunoglobulins have four peptide chains and Y-shaped structures, the TCR consists of just two peptide chains (α and β chains), both of which span the cytoplasmic membrane of the T cell.
TCRs are epitope-specific, and it has been estimated that 25 million T cells with unique epitope-binding TCRs are required to protect an individual against a wide range of microbial pathogens. Because the human genome only contains about 25,000 genes, we know that each specific TCR cannot be encoded by its own set of genes. This raises the question of how such a vast population of T cells with millions of specific TCRs can be achieved. The answer is a process called genetic rearrangement, which occurs in the thymus during the first step of thymic selection.
The genes that code for the variable regions of the TCR are divided into distinct gene segments called variable (V), diversity (D), and joining (J) segments. The genes segments associated with the α chain of the TCR consist 70 or more different Vα segments and 61 different Jα segments. The gene segments associated with the β chain of the TCR consist of 52 different Vβ segments, two different Dβ segments, and 13 different Jβ segments. During the development of the functional TCR in the thymus, genetic rearrangement in a T cell brings together one Vα segment and one Jα segment to code for the variable region of the α chain. Similarly, genetic rearrangement brings one of the Vβ segments together with one of the Dβ segments and one of thetJβ segments to code for the variable region of the β chain. All the possible combinations of rearrangements between different segments of V, D, and J provide the genetic diversity required to produce millions of TCRs with unique epitope-specific variable regions.
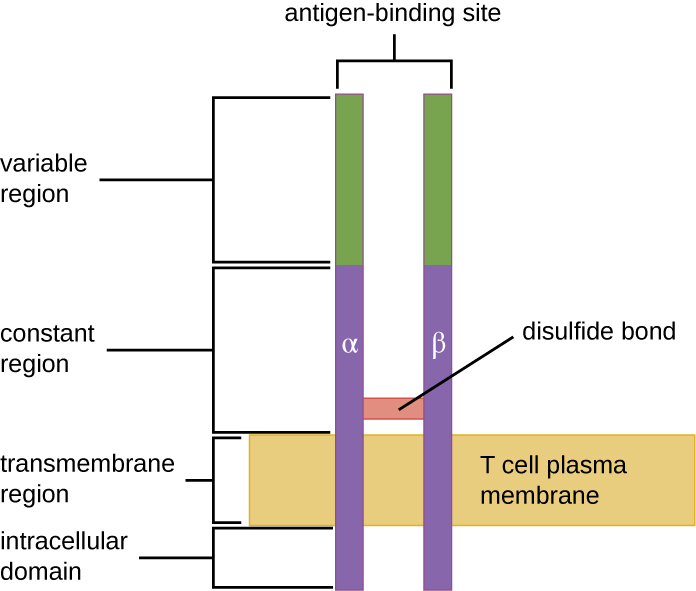
- What are the similarities and differences between TCRs and immunoglobulins?
- What process is used to provide millions of unique TCR binding sites?
Activation and Differentiation of Helper T Cells
Helper T cells can only be activated by APCs presenting processed foreign epitopes in association with MHC II. The first step in the activation process is TCR recognition of the specific foreign epitope presented within the MHC II antigen-binding cleft. The second step involves the interaction of CD4 on the helper T cell with a region of the MHC II molecule separate from the antigen-binding cleft. This second interaction anchors the MHC II-TCR complex and ensures that the helper T cell is recognizing both the foreign (“nonself”) epitope and “self” antigen of the APC; both recognitions are required for activation of the cell. In the third step, the APC and T cell secrete cytokines that activate the helper T cell. The activated helper T cell then proliferates, dividing by mitosis to produce clonal naïve helper T cells that differentiate into subtypes with different functions (Figure 19.17).
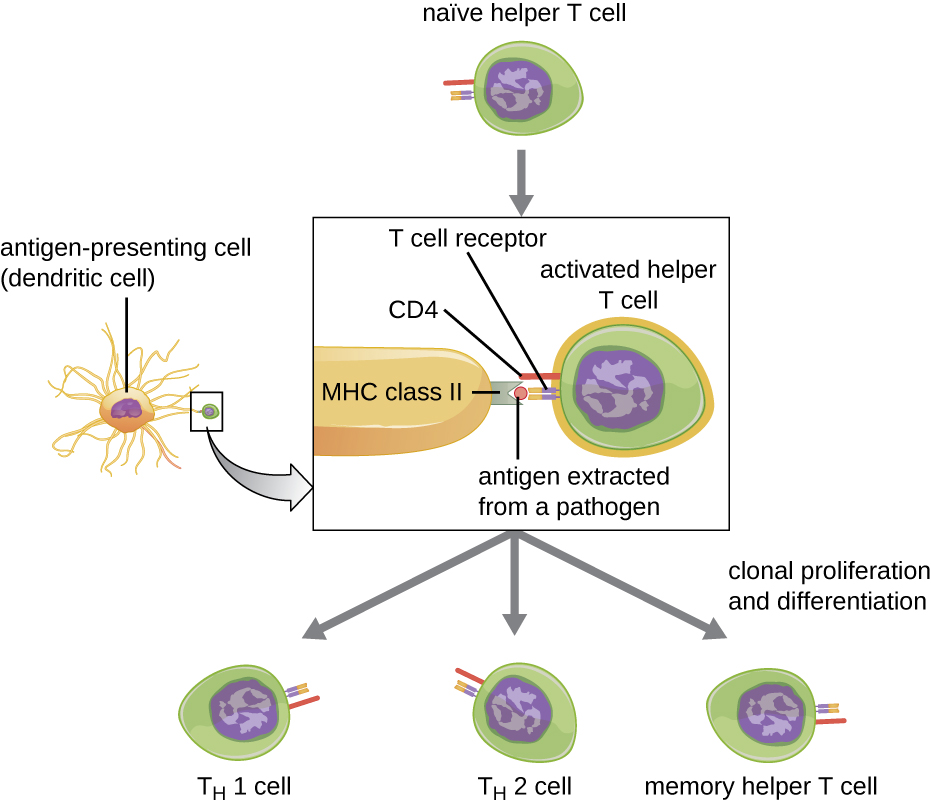
Activated helper T cells can differentiate into one of four distinct subtypes, summarized in Table 19.2. The differentiation process is directed by APC-secreted cytokines. Depending on which APC-secreted cytokines interact with an activated helper T cell, the cell may differentiate into a T helper 1 (TH1) cell, a T helper 2 (TH2) cell, or a memory helper T cell. The two types of helper T cells are relatively short-lived effector cells, meaning that they perform various functions of the immediate immune response. In contrast, memory helper T cells are relatively long lived; they are programmed to “remember” a specific antigen or epitope in order to mount a rapid, strong, secondary response to subsequent exposures.
TH1 cells secrete their own cytokines that are involved in stimulating and orchestrating other cells involved in adaptive and innate immunity. For example, they stimulate cytotoxic T cells, enhancing their killing of infected cells and promoting differentiation into memory cytotoxic T cells. TH1 cells also stimulate macrophages and neutrophils to become more effective in their killing of intracellular bacteria. They can also stimulate NK cells to become more effective at killing target cells.
TH2 cells play an important role in orchestrating the humoral immune response through their secretion of cytokines that activate B cells and direct B cell differentiation and antibody production. Various cytokines produced by TH2 cells orchestrate antibody class switching, which allows B cells to switch between the production of IgM, IgG, IgA, and IgE as needed to carry out specific antibody functions and to provide pathogen-specific humoral immune responses.
A third subtype of helper T cells called TH17 cells was discovered through observations that immunity to some infections is not associated with TH1 or TH2 cells. TH17 cells and the cytokines they produce appear to be specifically responsible for the body’s defence against chronic mucocutaneous infections. Patients who lack sufficient TH17 cells in the mucosa (e.g., HIV patients) may be more susceptible to bacteraemia and gastrointestinal infections.[1]
Table 19.2. Subtypes of Helper T Cells
| Subtypes of Helper T Cells | |
|---|---|
| Subtype | Functions |
| TH1 cells | Stimulate cytotoxic T cells and produce memory cytotoxic T cells |
| Stimulate macrophages and neutrophils (PMNs) for more effective intracellular killing of pathogens | |
| Stimulate NK cells to kill more effectively | |
| TH2 cells | Stimulate B cell activation and differentiation into plasma cells and memory B cells |
| Direct antibody class switching in B cells | |
| TH17 cells | Stimulate immunity to specific infections such as chronic mucocutaneous infections |
| Memory helper T cells | “Remember” a specific pathogen and mount a strong, rapid secondary response upon re-exposure |
Activation and Differentiation of Cytotoxic T Cells
Cytotoxic T cells (also referred to as cytotoxic T lymphocytes, or CTLs) are activated by APCs in a three-step process similar to that of helper T cells. The key difference is that the activation of cytotoxic T cells involves recognition of an antigen presented with MHC I (as opposed to MHC II) and interaction of CD8 (as opposed to CD4) with the receptor complex. After the successful co-recognition of foreign epitope and self-antigen, the production of cytokines by the APC and the cytotoxic T cell activate clonal proliferation and differentiation. Activated cytotoxic T cells can differentiate into effector cytotoxic T cells that target pathogens for destruction or memory cells that are ready to respond to subsequent exposures.
As noted, proliferation and differentiation of cytotoxic T cells is also stimulated by cytokines secreted from TH1 cells activated by the same foreign epitope. The co-stimulation that comes from these TH1 cells is provided by secreted cytokines. Although it is possible for activation of cytotoxic T cells to occur without stimulation from TH1 cells, the activation is not as effective or long-lasting.
Once activated, cytotoxic T cells serve as the effector cells of cellular immunity, recognizing and kill cells infected with intracellular pathogens through a mechanism very similar to that of NK cells. However, whereas NK cells recognize nonspecific signals of cell stress or abnormality, cytotoxic T cells recognize infected cells through antigen presentation of pathogen-specific epitopes associated with MHC I. Once an infected cell is recognized, the TCR of the cytotoxic T cell binds to the epitope and releases perforin and granzymes that destroy the infected cell (Figure 19.18). Perforin is a protein that creates pores in the target cell, and granzymes are proteases that enter the pores and induce apoptosis. This mechanism of programmed cell death is a controlled and efficient means of destroying and removing infected cells without releasing the pathogens inside to infect neighbouring cells, as might occur if the infected cells were simply lysed.
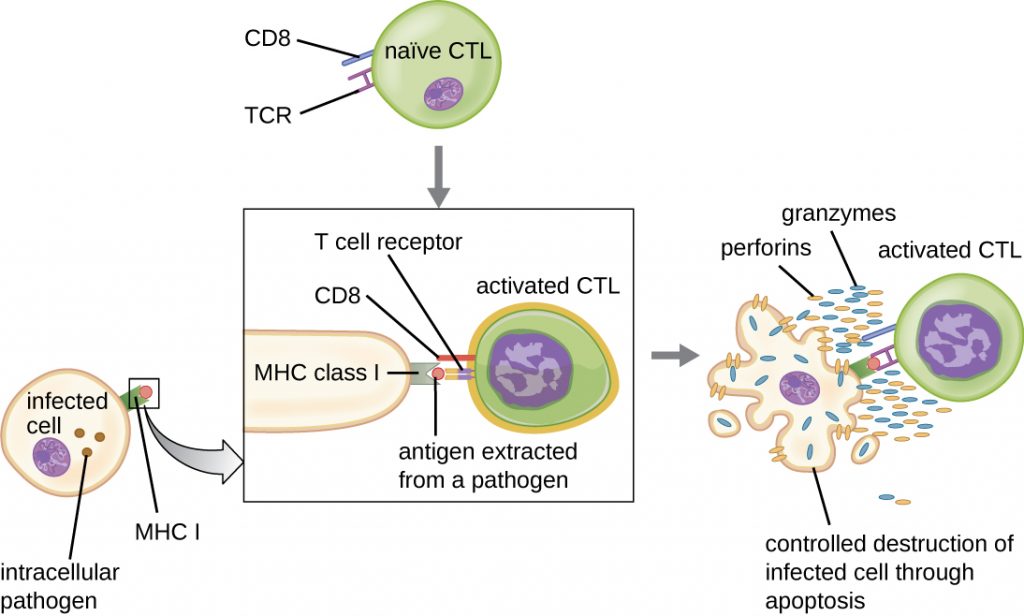

- Compare and contrast the activation of helper T cells and cytotoxic T cells.
- What are the different functions of helper T cell subtypes?
- What is the mechanism of CTL-mediated destruction of infected cells?
Superantigens and Unregulated Activation of T Cells
When T cell activation is controlled and regulated, the result is a protective response that is effective in combating infections. However, if T cell activation is unregulated and excessive, the result can be a life-threatening. Certain bacterial and viral pathogens produce toxins known as superantigens (see Virulence Factors of Bacterial and Viral Pathogens) that can trigger such an unregulated response. Known bacterial superantigens include toxic shock syndrome toxin (TSST), staphylococcal enterotoxins, streptococcal pyrogenic toxins, streptococcal superantigen, and the streptococcal mitogenic exotoxin. Viruses known to produce superantigens include Epstein-Barr virus (human herpesvirus 4), cytomegalovirus (human herpesvirus 5), and others.
The mechanism of T cell activation by superantigens involves their simultaneous binding to MHC II molecules of APCs and the variable region of the TCR β chain. This binding occurs outside of the antigen-binding cleft of MHC II, so the superantigen will bridge together and activate MHC II and TCR without specific foreign epitope recognition (Figure 19.19). The result is an excessive, uncontrolled release of cytokines, often called a cytokine storm, which stimulates an excessive inflammatory response. This can lead to a dangerous decrease in blood pressure, shock, multi-organ failure, and potentially, death.
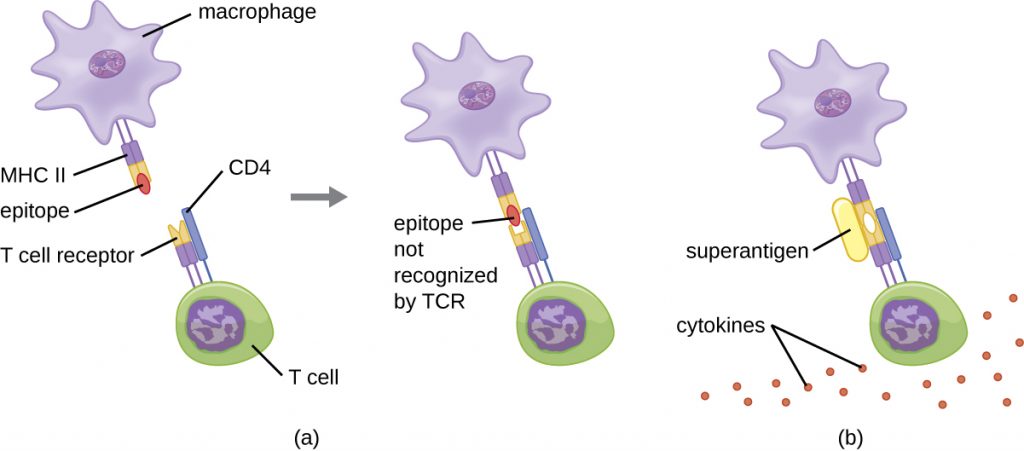
- What are examples of superantigens?
- How does a superantigen activate a helper T cell?
- What effect does a superantigen have on a T cell?
CASE IN POINT: Superantigens
Melissa, an otherwise healthy 22-year-old woman, is brought to the emergency room by her concerned boyfriend. She complains of a sudden onset of high fever, vomiting, diarrhoea, and muscle aches. In her initial interview, she tells the attending physician that she is on hormonal birth control and also is two days into the menstruation portion of her cycle. She is on no other medications and is not abusing any drugs or alcohol. She is not a smoker. She is not diabetic and does not currently have an infection of any kind to her knowledge.
While waiting in the emergency room, Melissa’s blood pressure begins to drop dramatically and her mental state deteriorates to general confusion. The physician believes she is likely suffering from toxic shock syndrome (TSS). TSS is caused by the toxin TSST-1, a superantigen associated with Staphylococcus aureus, and improper tampon use is a common cause of infections leading to TSS. The superantigen inappropriately stimulates widespread T cell activation and excessive cytokine release, resulting in a massive and systemic inflammatory response that can be fatal.
Vaginal or cervical swabs may be taken to confirm the presence of the microbe, but these tests are not critical to perform based on Melissa’s symptoms and medical history. The physician prescribes rehydration, supportive therapy, and antibiotics to stem the bacterial infection. She also prescribes drugs to increase Melissa’s blood pressure. Melissa spends three days in the hospital undergoing treatment; in addition, her kidney function is monitored because of the high risk of kidney failure associated with TSS. After 72 hours, Melissa is well enough to be discharged to continue her recovery at home.
- In what way would antibiotic therapy help to combat a superantigen?
CLINICAL FOCUS: Part 2
Olivia’s swollen lymph nodes, abdomen, and spleen suggest a strong immune response to a systemic infection in progress. In addition, little Olivia is reluctant to turn her head and appears to be experiencing severe neck pain. The physician orders a complete blood count, blood culture, and lumbar puncture. The cerebrospinal fluid (CSF) obtained appears cloudy and is further evaluated by Gram stain assessment and culturing for potential bacterial pathogens. The complete blood count indicates elevated numbers of white blood cells in Olivia’s bloodstream. The white blood cell increases are recorded at 28.5 K/µL (normal range: 6.0–17.5 K/µL). The neutrophil percentage was recorded as 60% (normal range: 23–45%). Glucose levels in the CSF were registered at 30 mg/100 mL (normal range: 50–80 mg/100 mL). The WBC count in the CSF was 1,163/mm3 (normal range: 5–20/mm3).
- Based on these results, do you have a preliminary diagnosis?
- What is a recommended treatment based on this preliminary diagnosis?
Jump to the next Clinical Focus box. Go back to the previous Clinical Focus box.
Key Takeaways
- Immature T lymphocytes are produced in the red bone marrow and travel to the thymus for maturation.
- Thymic selection is a three-step process of negative and positive selection that determines which T cells will mature and exit the thymus into the peripheral bloodstream.
- Central tolerance involves negative selection of self-reactive T cells in the thymus, and peripheral tolerance involves anergy and regulatory T cells that prevent self-reactive immune responses and autoimmunity.
- The TCR is similar in structure to immunoglobulins, but less complex. Millions of unique epitope-binding TCRs are encoded through a process of genetic rearrangement of V, D, and J gene segments.
- T cells can be divided into three classes—helper T cells, cytotoxic T cells, and regulatory T cells—based on their expression of CD4 or CD8, the MHC molecules with which they interact for activation, and their respective functions.
- Activated helper T cells differentiate into TH1, TH2, TH17, or memory T cell subtypes. Differentiation is directed by the specific cytokines to which they are exposed. TH1, TH2, and TH17 perform different functions related to stimulation of adaptive and innate immune defences. Memory T cells are long-lived cells that can respond quickly to secondary exposures.
- Once activated, cytotoxic T cells target and kill cells infected with intracellular pathogens. Killing requires recognition of specific pathogen epitopes presented on the cell surface using MHC I molecules. Killing is mediated by perforin and granzymes that induce apoptosis.
- Superantigens are bacterial or viral proteins that cause a nonspecific activation of helper T cells, leading to an excessive release of cytokines (cytokine storm) and a systemic, potentially fatal inflammatory response.
Multiple Choice
Fill in the Blank
Short Answer
- What is the basic difference in effector function between helper and cytotoxic T cells?
- What necessary interactions are required for activation of helper T cells and activation/effector function of cytotoxic T cells?
Media Attributions
- Figure_06_03_01
- OSC_Microbio_18_01_marrow
- OSC_Microbio_18_01_thymus
- OSC_Microbio_18_03_TCR
- OSC_Microbio_18_03_Tcellact
- OSC_Microbio_18_03_Tprolif
- microbiology sign © Nick Youngson
- OSC_Microbio_18_03_SuperAntig
- Blaschitz C., Raffatellu M. “Th17 cytokines and the gut mucosal barrier.” J Clin Immunol. 2010 Mar; 30(2):196-203. doi: 10.1007/s10875-010-9368-7. ↵
