7.6 Liens entre les voies métaboliques des glucides, des protéines et des lipides
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Discuter des liens entre les voies métaboliques des glucides, la glycolyse et le cycle de l’acide citrique avec les voies métaboliques des protéines et des lipides
- Expliquer pourquoi les voies métaboliques ne sont pas considérées comme des systèmes fermés
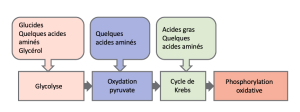
Vous avez appris le catabolisme du glucose, qui fournit de l’énergie aux cellules vivantes. Mais les êtres vivants consomment des composés organiques autres que le glucose pour se nourrir. Comment un sandwich à la dinde finira-t-il par être de l’ATP dans vos cellules ? Cela se produit parce que toutes les voies cataboliques des glucides, des protéines et des lipides finissent par se relier à la glycolyse et aux voies du cycle de l’acide citrique (voir la Figure 7.19). Les voies métaboliques doivent être considérées comme poreuses et interconnectées, c’est-à-dire que les substances entrent par d’autres voies et que les intermédiaires partent pour d’autres voies. Ces voies ne sont pas des systèmes fermés ! Bon nombre des substrats, intermédiaires et produits d’une voie particulière sont des réactifs dans d’autres voies.
Liens entre les autres sucres et le métabolisme du glucose
Le glycogène, un polymère du glucose, est une molécule de stockage d’énergie chez les animaux. Lorsqu’il y a suffisamment d’ATP, l’excès de glucose est stocké sous forme de glycogène dans les cellules hépatiques et musculaires. Le glycogène sera hydrolysé en monomères de glucose 1-phosphate (G-1-P) si le taux de sucre dans le sang baisse. La présence de glycogène comme source de glucose permet de produire de l’ATP pendant une plus longue période pendant l’exercice. Le glycogène est décomposé en glucose-1-phosphate (G-1-P) et converti en glucose-6-phosphate (G-6-P) dans les cellules musculaires et hépatiques, et ce produit entre dans la voie glycolytique.
Le saccharose est un disaccharide avec une molécule de glucose et une molécule de fructose liées ensemble par une liaison glycosidique. Le fructose est l’un des trois monosaccharides « alimentaires », ainsi que le glucose et le galactose (qui font partie du lactose disaccharide de sucre du lait), qui sont absorbés directement dans la circulation sanguine pendant la digestion. Le catabolisme du fructose et du galactose produit le même nombre de molécules d’ATP que le glucose.
Liens entre les autres sucres et le métabolisme du glucose
Les protéines sont hydrolysées par une variété d’enzymes dans les cellules. La plupart du temps, les acides aminés sont recyclés dans la synthèse de nouvelles protéines. Toutefois, s’il y a un excès d’acides aminés ou si le corps est dans un état de famine, certains acides aminés seront mis en dérivation dans les voies du catabolisme du glucose (Figure 7.18). Il est très important de noter que chaque acide aminé doit voir son groupement aminé retiré avant d’entrer dans ces voies. Le groupement amino est converti en ammoniac. Chez les mammifères, le foie synthétise l’urée à partir de deux molécules d’ammoniac et d’une molécule de dioxyde de carbone. Ainsi, l’urée est le principal déchet chez les mammifères, produit à partir de l’azote provenant des acides aminés, et elle quitte le corps dans l’urine. Il est à noter que les acides aminés peuvent être synthétisés à partir des intermédiaires et des réactifs du cycle de respiration cellulaire.

Liens entre les métabolismes des lipides et du glucose
Les lipides reliés à la voie du glucose comprennent le cholestérol et les triglycérides. Le cholestérol est un lipide qui contribue à la flexibilité des membranes cellulaires et est un précurseur des hormones stéroïdiennes. La synthèse du cholestérol commence par les groupements acétyles et se poursuit dans une seule direction. Le processus ne peut pas être inversé.
Les triglycérides, fabriqués à partir de la liaison du glycérol et de trois acides gras, sont une forme de stockage d’énergie à long terme chez les animaux. Les animaux peuvent produire la plus grande partie des acides gras dont ils ont besoin. Les triglycérides peuvent être fabriqués et décomposés par certaines parties des voies de catabolisme du glucose. Le glycérol peut être phosphorylé en glycérol-3-phosphate, qui se poursuit par glycolyse. Les acides gras sont catabolisés dans un processus appelé bêta-oxydation, qui se produit dans la matrice des mitochondries et convertit leurs chaînes d’acides gras en unités à deux carbones de groupements acétyle. Les groupements acétyle sont ramassés par la CoA pour former de l’acétyl CoA qui passe dans le cycle de l’acide citrique.
Lien avec l’évolution
Voies de photosynthèse et métabolisme cellulaire
Les processus de photosynthèse et de métabolisme cellulaire consistent en plusieurs voies très complexes. On pense généralement que les premières cellules sont apparues dans un environnement aqueux — une « soupe » de nutriments — peut-être à la surface de certaines argiles poreuses, peut-être dans des milieux marins chauds. Si ces cellules se reproduisent avec succès et que leur nombre augmente de façon constante, il s’ensuit que les cellules commenceraient à appauvrir les éléments nutritifs du milieu dans lequel elles vivaient à mesure qu’elles transforment les éléments nutritifs en composants de leur propre corps. Cette situation hypothétique aurait entraîné une sélection naturelle favorisant les organismes qui pourraient exister en utilisant les nutriments qui restent dans leur environnement et en manipulant ces éléments nutritifs en matériaux avec lesquels ils pourraient survivre. La sélection favoriserait les organismes qui pourraient extraire la valeur maximale des nutriments auxquels ils avaient accès.
Une première forme de photosynthèse s’est développée qui exploitait l’énergie du soleil en utilisant l’eau comme source d’atomes d’hydrogène, mais cette voie ne produisait pas d’oxygène libre (photosynthèse anoxygène). (Un autre type de photosynthèse anoxygène ne produisait pas d’oxygène libre parce qu’il n’utilisait pas l’eau comme source d’ions hydrogène ; il utilisait plutôt des matériaux comme le sulfure d’hydrogène et, par conséquent, du soufre.) On pense que la glycolyse s’est développée à ce moment-là et qu’elle pourrait tirer profit des sucres simples produits, mais que ces réactions n’ont pas permis d’extraire complètement l’énergie emmagasinée dans les glucides. L’apparition de la glycolyse était probablement antérieure à l’évolution de la photosynthèse, car elle était bien adaptée pour extraire l’énergie des matières qui s’accumulaient spontanément dans la « soupe primaire ». Une forme ultérieure de photosynthèse utilisait l’eau comme source d’électrons et d’hydrogène et produisait de l’oxygène libre. Au fil du temps, l’atmosphère s’est oxygénée, mais pas avant que l’oxygène libère des métaux oxydés dans l’océan et crée une couche de « rouille » dans les sédiments, ce qui a permis de dater l’apparition des premiers photosynthétiseurs oxygénés. Les êtres vivants se sont adaptés pour exploiter cette nouvelle atmosphère qui a permis à la respiration aérobie telle que nous la connaissons d’évoluer. Lorsque le processus complet de photosynthèse oxygénique s’est développé et que l’atmosphère est devenue oxygénée, les cellules ont finalement pu utiliser l’oxygène expulsé par photosynthèse pour extraire beaucoup plus d’énergie des molécules de sucre en utilisant le cycle de l’acide citrique et la phosphorylation oxydative.
