10.1 La division cellulaire
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de :
- Décrire la structure des génomes procaryotes et eucaryotes
- Faire la distinction entre les chromosomes, les gènes et les caractères
- Décrire les mécanismes de la compaction chromosomique
La reproduction des cellules, grâce au cycle cellulaire, est à la base même de la continuité de la vie. Le cycle cellulaire est une séquence ordonnée d’événements qui décrit les étapes de la vie d’une cellule, de la division d’une cellule mère à la production de deux cellules filles génétiquement identiques.
ADN génomique
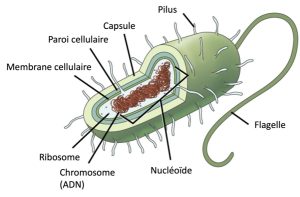
Avant de discuter des étapes qu’une cellule doit suivre pour répliquer et diviser son ADN, il est nécessaire de bien comprendre la structure et la fonction de l’information génétique d’une cellule. Sous la forme d’une molécule d’ADN à double brin, l’ADN d’une cellule est son génome. Chez les procaryotes, le génome est composé d’une simple molécule d’ADN à double brin qui prend la forme d’une boucle ou d’un cercle (Figure 10.2) Le nucléoïde est la région de la cellule qui contient ce matériel génétique. Certains procaryotes ont également de plus petites boucles d’ADN appelées plasmides qui ne sont pas essentiels à une croissance normale. Les bactéries peuvent échanger leurs plasmides avec d’autres bactéries, et parfois recevoir de nouveaux gènes bénéfiques qu’elles peuvent ajouter à leur ADN chromosomique. La résistance aux antibiotiques est un trait qui se répand souvent au sein d’une colonie bactérienne grâce à l’échange de plasmides entre des donneurs résistants vers des cellules receveuses.
Chez les eucaryotes, le génome est constitué de plusieurs molécules d’ADN à double brin linéaires (Figure 10.3). Chaque espèce d’eucaryotes possède un nombre caractéristique de chromosomes dans le noyau de ses cellules. Les cellules du corps humain (cellules somatiques) ont 46 chromosomes, alors que les gamètes humains (spermatozoïdes ou ovules) n’en ont que 23 chacun. Une cellule somatique typique contient deux jeux de chromosomes appariés ou homologues (un jeu provenant de chaque parent biologique) – une configuration que l’on appelle diploïde. (Nota : La lettre n est utilisée pour représenter un jeu de chromosomes; par conséquent, un organisme diploïde est désigné sous le terme 2n.) Les cellules humaines qui contiennent un jeu de chromosomes sont appelées des gamètes, ou des cellules sexuelles; il s’agit des ovules et des spermatozoïdes, que l’on désigne par cellules 1n ou haploïdes.
Au moment de la fécondation, chaque gamète apporte un jeu de chromosomes, créant ainsi une cellule diploïde qui contient des paires de chromosomes appelés chromosomes homologues (« qui correspondent à »). Les chromosomes homologues sont de la même longueur et ont des segments de nucléotides spécifiques, appelés gènes, situés exactement au même endroit, ou locus. Les gènes, c’est-à-dire les unités fonctionnelles des chromosomes, déterminent des caractères particuliers en codant pour des protéines spécifiques. Les traits sont des variations de ces caractères. Par exemple, la couleur des cheveux est un caractère, et les traits correspondront aux cheveux blonds, bruns, noirs, et toutes les nuances de ces couleurs.
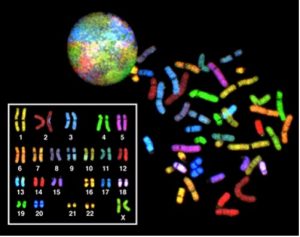
Chaque copie d’une paire de chromosomes homologues provient d’un parent différent; par conséquent, les différents gènes (allèles) ne sont pas identiques, bien qu’ils codent pour les mêmes traits, par exemple la « couleur des cheveux ». Les différences entre les individus d’une même espèce résultent de la combinaison de gènes hérités des deux parents. Même une légère différence dans la séquence des nucléotides d’un gène peut produire un trait différent. Par exemple, il existe trois séquences génétiques possibles sur le chromosome humain qui portent le code pour le groupe sanguin : la séquence A, la séquence B et la séquence O. Sachant que toutes les cellules humaines diploïdes possèdent deux copies du chromosome qui détermine le groupe sanguin, le groupe sanguin (le trait) est déterminé par les deux allèles du gène marqueur qui sont hérités. Il est possible d’avoir deux copies de la même séquence génétique sur les deux chromosomes homologues, avec une sur chacun d’entre eux (par exemple, AA, BB ou OO), ou deux séquences différentes, comme AB, AO ou BO.
Apparemment, les variations mineures de traits, telles que le groupe sanguin, la couleur des yeux et la chiralité, contribuent à la variation naturelle qui existe au sein d’une espèce, mais même s’ils semblent mineurs, ces traits peuvent être connectés à l’expression d’autres traits encore inconnus. Toutefois, si la séquence entière d’ADN d’une paire quelconque de chromosomes homologues humains est comparée, la différence est bien inférieure à un pour cent. Les chromosomes sexuels, X et Y, sont la seule exception à la règle de l’uniformité des chromosomes homologues : À part un faible pourcentage d’homologie nécessaire pour produire des gamètes avec précision, les gènes que l’on retrouve sur les chromosomes X et Y sont différents.
Structure et compaction des chromosomes chez les eucaryotes
Si l’ADN de tous les 46 chromosomes d’un noyau de cellule humaine était étalé bout à bout, il mesurerait environ deux mètres; toutefois, son diamètre ne serait que de 2 nm ! Sachant que la taille d’une cellule humaine typique est d’environ 10 µm (100 000 cellules alignées bout à bout pour faire un mètre), l’ADN doit être empaqueté de façon serrée pour pouvoir rentrer dans le noyau d’une cellule. Mais il doit en même temps être facilement accessible pour permettre l’expression des gènes. C’est pour cette raison que les longs brins d’ADN sont condensés dans des chromosomes compacts durant certaines étapes du cycle cellulaire. La compaction des chromosomes se fait de plusieurs façons.
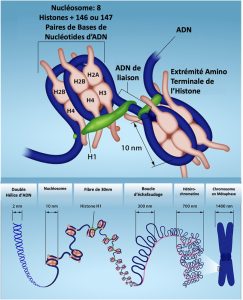
Au premier niveau de la compaction, les fragments courts de la double hélice d’ADN s’enroulent autour d’un cœur formé de huit histones, à des intervalles réguliers sur toute la longueur du chromosome (Figure 10.4). Le complexe ADN-histones s’appelle la chromatine. L’unité ADN-histone en forme de perle s’appelle le nucléosome, et l’ADN qui relie les nucléosomes entre eux s’appelle l’ADN lieur. Une molécule d’ADN sous cette forme est environ sept fois plus courte qu’une double hélice sans histones, et les perles mesurent environ 10 nm de diamètre, par contraste avec le diamètre de 2 nm d’une double hélice d’ADN.
Le second niveau de compaction survient lorsque les nucléosomes et l’ADN lieur qui les sépare s’enroulent eux aussi en une fibre de chromatine de 30 nm. Cette spirale condense encore davantage le chromosome qui est maintenant 50 fois plus court que sous sa forme étendue.
Au troisième niveau de compaction, diverses protéines fibreuses sont utilisées pour « empaqueter la chromatine ». Ces protéines fibreuses veillent également à ce que chaque chromosome d’une cellule non proliférative occupe une région particulière du noyau qui ne chevauche pas celle d’un autre chromosome (voir l’image de la partie supérieure de la Figure 10.3).
La réplication de l’ADN survient dans la phase S de l’interphase, qui ne fait techniquement pas partie de la mitose, mais qui la précède toujours. Après la réplication, les chromosomes sont composés de deux chromatides sœurs accolées. Lorsqu’elles sont complètement compactes, les paires de chromosomes empaquetés de façon identique sont liées entre elles au moyen des protéines cohésines. Le centromère est la région où la liaison entre les chromatides sœurs est la plus étroite. Les chromatides sœurs accolées, dont le diamètre est d’environ 1 µm, sont visibles grâce à un microscope optique. La région centromérique est hautement condensée et apparaîtra donc comme une zone étranglée.
Lien vers l’apprentissage
Cette animation illustre les différents niveaux de l’empaquetage chromosomique.
Cliquez pour voir le contenu (https://www.openstax.org/l/Packaged_DNA) (en anglais)
