5.1 Composants et structure
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Comprendre le modèle de mosaïque des fluides membranaires cellulaires
- Décrire les fonctions phospholipides, protéiques et glucidiques dans les membranes
- Discuter de la fluidité de la membrane
La membrane plasmique d’une cellule définit la cellule, décrit ses frontières et détermine la nature de son interaction avec son environnement (voir le Tableau 5.1 pour un résumé). Les cellules excluent certaines substances, en prennent d’autres et en excrètent encore d’autres, le tout en quantités contrôlées. La membrane plasmique doit être très souple pour permettre à certaines cellules, comme les globules rouges et les globules blancs, de changer de forme lorsqu’elles traversent des capillaires étroits. Ce sont les fonctions les plus évidentes de la membrane plasmique. De plus, la surface de la membrane plasmique porte des marqueurs qui permettent aux cellules de se reconnaître mutuellement, ce qui est essentiel à la formation des tissus et des organes au début du développement, et qui joue plus tard un rôle dans la distinction entre le « soi » et le « non-soi » de la réponse immunitaire.
Parmi les fonctions les plus sophistiquées de la membrane plasmique, on compte la capacité des récepteurs de protéines intégrales complexes à transmettre des signaux. Ces protéines agissent à la fois comme récepteurs d’entrée extracellulaires et comme activateurs de traitement intracellulaire. Ces récepteurs membranaires fournissent des sites de fixation extracellulaires pour des effecteurs comme les hormones et les facteurs de croissance, et ils activent les cascades de réponse intracellulaire lorsque leurs effecteurs sont liés. Parfois, les virus détournent des récepteurs (VIH, virus de l’immunodéficience humaine, en est un exemple) qui les utilisent pour entrer dans les cellules, et parfois, les gènes codant les récepteurs deviennent mutés, ce qui entraîne un dysfonctionnement du processus de transduction du signal avec des conséquences désastreuses.
Modèle de la mosaïque fluide
Les scientifiques ont identifié la membrane plasmique dans les années 1890 et ses composants chimiques en 1915. Les principaux composants qu’ils ont identifiés étaient les lipides et les protéines. En 1935, Hugh Davson et James Danielli proposent la structure de la membrane plasmique. Il s’agit du premier modèle que d’autres membres de la communauté scientifique ont largement accepté. Elle était fondée sur l’aspect « voie ferrée » de la membrane plasmique dans les premières micrographies électroniques. Davson et Danielli ont émis l’hypothèse que la structure de la membrane plasmique ressemble à un sandwich. Ils ont fait l’analogie entre les protéines et le pain, et les lipides à la garniture. Dans les années 1950, les progrès de la microscopie, notamment la microscopie électronique à transmission (MET), ont permis aux chercheurs de voir que le noyau de la membrane plasmique était constitué d’une double couche plutôt que d’une seule couche. En 1972, S.J. Singer et Garth L. Nicolson ont proposé un nouveau modèle qui fournit des observations microscopiques et explique mieux la fonction de la membrane plasmique.
L’explication, le modèle de la mosaïque fluide, a quelque peu évolué au fil du temps, mais elle tient compte encore mieux de la structure et de la fonction de la membrane plasmique telle que nous la comprenons maintenant. Le modèle de la mosaïque fluide décrit la structure de la membrane plasmique comme une mosaïque de composants — y compris les phospholipides, le cholestérol, les protéines et les hydrates de carbone — qui confèrent à la membrane un caractère fluide. Les membranes du plasma varient de 5 à 10 nm d’épaisseur. À titre de comparaison, les globules rouges humains, visibles par microscopie optique, mesurent environ 8 µm de largeur, soit environ 1 000 fois plus larges qu’une membrane plasmique. La membrane ressemble un peu à un sandwich (Figure 5.2).
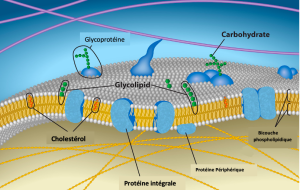
Les principaux composants d’une membrane plasmique sont les lipides (phospholipides et cholestérol), les protéines et les glucides attachés à certains lipides et protéines. Un phospholipide est une molécule constituée de glycérol, de deux acides gras et d’un groupe de tête lié au phosphate. Le cholestérol, un autre lipide composé de quatre cycles de carbone fondu, est situé le long des phospholipides au cœur de la membrane. Les proportions de protéines, de lipides et de glucides dans la membrane plasmique varient selon le type de cellule, mais pour une cellule humaine typique, les protéines représentent environ 50 % de la composition en masse, les lipides (de tous les types) représentent environ 40 % et les glucides représentent les 10 % restants. Cependant, la concentration en protéines et en lipides varie selon les membranes cellulaires. Par exemple, la myéline, une excroissance de la membrane des cellules spécialisées qui isole les axones des nerfs périphériques, ne contient que 18 % de protéines et 76 % de lipides. La membrane interne mitochondriale contient 76 % de protéines et seulement 24 % de lipides. La membrane plasmique des globules rouges humains est constituée de 30 % de lipides. Les glucides ne sont présents que sur la surface extérieure de la membrane plasmique et sont attachés aux protéines, formant des glycoprotéines ou s’ils sont attachés aux lipides, formant des glycolipides.
Phospholipides
Le tissu principal de la membrane comprend des molécules amphiphiles phospholipides. Les zones hydrophiles ou « aimantes d’eau » de ces molécules (qui ressemblent à une collection de boules dans l’interprétation du modèle par un artiste) (Figure 5.2) sont en contact avec le fluide aqueux à l’intérieur et à l’extérieur de la cellule. Les molécules hydrophobes, ou qui détestent l’eau, ont tendance à être non polaires. Ils interagissent avec d’autres molécules non polaires lors de réactions chimiques, mais n’interagissent généralement pas avec les molécules polaires. Lorsqu’elles sont placées dans l’eau, les molécules hydrophobes ont tendance à former une boule ou une grappe. Les régions hydrophiles des phospholipides forment des liaisons hydrogène avec l’eau et d’autres molécules polaires à l’extérieur et à l’intérieur de la cellule. Ainsi, les surfaces de la membrane qui font face à l’intérieur et à l’extérieur de la cellule sont hydrophiles. En revanche, l’intérieur de la membrane cellulaire est hydrophobe et n’interagit pas avec l’eau. Par conséquent, les phospholipides forment une excellente membrane cellulaire à deux couches qui sépare le fluide à l’intérieur de la cellule du fluide à l’extérieur de la cellule.
Une molécule phospholipidique (Figure 5.3) est constituée d’un squelette de glycérol à trois carbones avec deux molécules d’acides gras fixées aux carbones 1 et 2, et d’un groupe contenant du phosphate attaché au troisième carbone. Cette disposition donne à la molécule globale une zone de tête (le groupe contenant du phosphate), qui a un caractère polaire ou une charge négative, et une zone de queue (les acides gras), qui n’a aucune charge. La tête peut former des liaisons hydrogène, mais la queue ne peut pas. Les scientifiques appellent une molécule ayant une zone chargée positivement ou négativement et une zone non chargée, ou non polaire, amphiphile ou « à double amour ».
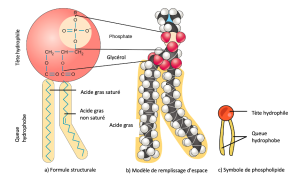
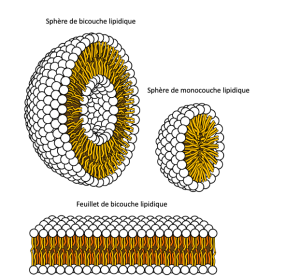
Cette caractéristique est essentielle à la structure de la membrane plasmique parce que, dans l’eau, les phospholipides s’organisent avec leurs queues hydrophobes face à face et leurs têtes hydrophiles tournées vers l’extérieur. De cette façon, ils forment une bicouche lipidique — une barrière phospholipidique double couche qui sépare l’eau et les autres matières d’un côté de l’eau et d’autres matériaux de l’autre côté. Les phospholipides chauffés dans une solution aqueuse forment habituellement spontanément de petites sphères ou gouttelettes (micelles ou liposomes), leurs têtes hydrophiles formant l’extérieur et leur queue hydrophobe à l’intérieur (Figure 5.4).
Protéines
Les protéines constituent le deuxième composant majeur des membranes plasmiques. Les protéines intégrales, ou intégrines, comme leur nom l’indique, s’intègrent complètement dans la structure membranaire, et leurs régions hydrophobes couvrant la membrane interagissent avec la région hydrophobe des bicouches phospholipides (Figure 5.2). Les protéines membranaires intégrales à passage unique ont habituellement un segment transmembranaire hydrophobe composé de 20 à 25 acides aminés. Certaines ne couvrent qu’une partie de la membrane, s’associant à une seule couche, tandis que d’autres s’étendent d’un côté à l’autre et sont exposées de chaque côté. Jusqu’à 12 segments protéiques uniques comprennent des protéines complexes, qui sont largement pliées et incorporées dans la membrane (Figure 5.5). Ce type de protéine a une ou plusieurs zones hydrophiles et une ou plusieurs zones légèrement hydrophobes. Cette disposition des zones protéiques oriente la protéine le long des phospholipides, la zone hydrophobe de la protéine étant adjacente aux queues des phospholipides et la ou les zones hydrophiles de la protéine dépassant de la membrane et en contact avec le cytosol ou le liquide extracellulaire.
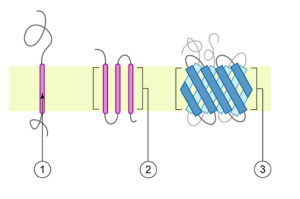
Les protéines périphériques se trouvent sur les surfaces extérieures et intérieures des membranes, fixées soit aux protéines intégrales, soit aux phospholipides. Les protéines périphériques, ainsi que les protéines intégrales, peuvent servir d’enzymes, d’attaches structurales pour les fibres du cytosquelette ou de sites de reconnaissance de la cellule. Les scientifiques appellent parfois ces protéines « spécifiques aux cellules ». L’organisme reconnaît ses propres protéines et attaque les protéines étrangères associées aux pathogènes invasifs.
Glucides
Les glucides sont le troisième composant principal de la membrane plasmique. Ils se trouvent toujours sur la surface extérieure des cellules et sont liés soit aux protéines (formant des glycoprotéines), soit aux lipides (formant des glycolipides) (Figure 5.2). Ces chaînes glucidiques peuvent être constituées de 2 à 60 unités de monosaccharide et peuvent être droites ou ramifiées. Avec les protéines périphériques, les glucides forment des sites spécialisés à la surface cellulaire qui permettent aux cellules de se reconnaître mutuellement. Ces sites ont des modèles uniques qui permettent la reconnaissance cellulaire, tout comme les traits faciaux propres à chaque personne permettent aux individus de la reconnaître. Cette fonction de reconnaissance est très importante pour les cellules, car elle permet au système immunitaire de faire la distinction entre les cellules corporelles (« soi ») et les cellules ou tissus étrangers (« non-soi »). Des types similaires de glycoprotéines et de glycolipides se trouvent à la surface des virus et peuvent changer fréquemment, ce qui empêche les cellules immunitaires de les reconnaître et de les attaquer.
Nous désignons collectivement ces glucides sur la surface extérieure de la cellule — les composants glucidiques des glycoprotéines et des glycolipides — sous le nom de glycocalyx (ce qui signifie « enrobage de sucre »). Le glycocalyx est très hydrophile et attire de grandes quantités d’eau à la surface de la cellule. Cela contribue à l’interaction de la cellule avec son environnement aqueux et à la capacité de la cellule à obtenir des substances dissoutes dans l’eau. Comme nous l’avons mentionné plus haut, le glycocalyx est également important pour l’identification cellulaire, l’autodétermination ou la non-autodétermination et le développement embryonnaire, et il est utilisé dans les attachements de cellules à cellules pour former des tissus.
Fluidité membranaire
La caractéristique de la mosaïque de la membrane aide à illustrer sa nature. Les protéines et les lipides intégraux existent dans la membrane sous forme de molécules séparées mais faiblement attachées. Ceux-ci ressemblent aux carreaux multicolores distincts d’une mosaïque, et ils flottent, se déplaçant quelque peu les uns par rapport aux autres. Cependant, la membrane n’est pas comme un ballon qui peut se dilater et se contracter ; elle est plutôt rigide et peut éclater si elle est pénétrée ou si une cellule reçoit trop d’eau. Cependant, en raison de sa nature mosaïque, une aiguille très fine peut facilement pénétrer une membrane plasmique sans provoquer son éclatement, et la membrane se referme lorsque l’on extrait l’aiguille.
Les caractéristiques de la mosaïque de la membrane expliquent une partie, mais pas la totalité de sa fluidité. Il y a deux autres facteurs qui aident à maintenir cette caractéristique de fluide. L’un des facteurs est la nature des phospholipides eux-mêmes. Sous leur forme saturée, les acides gras contenus dans les queues phospholipides sont saturés d’atomes d’hydrogène liés. Il n’y a pas de doubles liaisons entre les atomes de carbone adjacents. Il en résulte des queues relativement droites. En revanche, les acides gras insaturés ne contiennent pas un nombre maximal d’atomes d’hydrogène, mais ils contiennent des doubles liaisons entre les atomes de carbone adjacents. Une double liaison entraîne une courbure de la chaîne de carbone d’environ 30 degrés (Figure 5.3).
Ainsi, si des températures décroissantes compriment les acides gras saturés avec leur queue droite, ils se pressent les uns sur les autres, formant une membrane dense et assez rigide. Si les acides gras insaturés sont comprimés, les « plis » dans leur queue écartent les molécules phospholipides adjacentes, ce qui maintient un certain espace entre les molécules phospholipides. Cette « marge de manœuvre » aide à maintenir la fluidité de la membrane à des températures auxquelles les membranes contenant des résidus d’acides gras saturés dans leurs phospholipides « gèlent » ou se solidifient. La fluidité relative de la membrane est particulièrement importante dans un environnement froid. Un environnement froid comprime habituellement les membranes composées en grande partie d’acides gras saturés, ce qui les rend moins fluides et plus susceptibles de se rompre. De nombreux organismes (les poissons en sont un exemple) sont capables de s’adapter aux environnements froids en modifiant la proportion d’acides gras insaturés dans leurs membranes en réponse à une température plus basse.
Les animaux ont un composant membranaire supplémentaire qui aide à maintenir la fluidité. Le cholestérol, qui se trouve à côté des phospholipides dans la membrane, a tendance à atténuer les effets de la température sur la membrane. Ainsi, ce lipide fonctionne comme un tampon, empêchant les températures plus basses d’inhiber la fluidité et empêchant les températures accrues d’augmenter trop la fluidité. Ainsi, le cholestérol étend, dans les deux sens, la plage de température dans laquelle la membrane est convenablement fluide et, par conséquent, fonctionnelle. Le cholestérol remplit également d’autres fonctions, comme l’organisation de grappes de protéines transmembranaires en radeaux lipidiques.
Tableau 5.1 Composantes et fonctions de la membrane cellulaire
| Composante | Localisation |
| Phospholipide | Endroit centrale de la membrane |
| Cholestérol | Attaché entre phospholipides et entre les deux couches phospholipidiques |
| Protéines intégrales | Intégré dans les couches phospholipidique; ne doit pas pénétré les deux couches |
| Protéines périphériques | Sur la surface interne ou externe de la bicouche phospholipidique; n’est pas intégré dans les phospholipides |
| Glucides (composantes des glycoprotéines et glycolipides) | Généralement attaché à des protéines sur la surface externe de la membrane phospholipidique |
