16.2 Régulation des gènes chez les procaryotes
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Décrire les étapes de la régulation des gènes chez les procaryotes
- Expliquer les rôles des activateurs, des inducteurs et des répresseurs dans la régulation des gènes
L’ADN des procaryotes est organisé en un chromosome circulaire, condensé dans la région nucléoïde du cytoplasme de la cellule. Les protéines nécessaires à une fonction spécifique ou impliquées dans la même voie biochimique sont codées ensemble dans des blocs appelés opérons. Par exemple, tous les gènes nécessaires à l’utilisation du lactose comme source d’énergie sont codés les uns à côté des autres dans l’opéron lactose (ou lac) et transcrits dans un seul ARNm.
Dans les cellules procaryotes, il existe trois types de molécules régulatrices qui peuvent affecter l’expression des opérons : les répresseurs, les activateurs et les inducteurs. Les répresseurs et les activateurs sont des protéines produites dans la cellule. Les répresseurs et les activateurs régulent l’expression génique en se liant à des sites spécifiques de l’ADN adjacents aux gènes qu’ils contrôlent. En général, les activateurs se lient au site du promoteur, tandis que les répresseurs se lient aux régions de l’opérateur. Les répresseurs empêchent la transcription d’un gène en réponse à un stimulus externe, tandis que les activateurs augmentent la transcription d’un gène en réponse à un stimulus externe. Les inducteurs sont de petites molécules qui peuvent être produites par la cellule ou qui se trouvent dans l’environnement de la cellule. Les inducteurs activent ou répriment la transcription en fonction des besoins de la cellule et de la disponibilité du substrat.
L’opéron trp : Un opéron répressible
Les bactéries comme Escherichia coli ont besoin d’acides aminés pour survivre et sont capables d’en synthétiser un grand nombre. Le tryptophane est l’un de ces acides aminés qu’E. coli peut soit ingérer à partir de l’environnement, soit synthétiser à l’aide d’enzymes codées par cinq gènes. Ces cinq gènes se trouvent les uns à côté des autres dans ce que l’on appelle l’opéron tryptophane (trp) (figure 16.4). Les gènes sont transcrits en un seul ARNm, qui est ensuite traduit pour produire les cinq enzymes. Si le tryptophane est présent dans l’environnement, E. coli n’a pas besoin de le synthétiser et l’opéron trp est réprimé. Cependant, lorsque la disponibilité du tryptophane est faible, l’interrupteur contrôlant l’opéron est activé, l’ARNm est transcrit, les protéines enzymatiques sont traduites et le tryptophane est synthétisé.
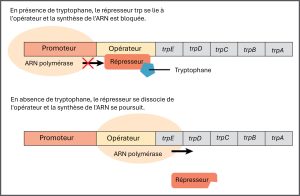
L’opéron trp comprend trois régions importantes : la région codante, l’opérateur trp et le promoteur trp. La région codante comprend les gènes des cinq enzymes de biosynthèse du tryptophane. Juste avant la région codante se trouve le site de départ de la transcription. La séquence promotrice, à laquelle l’ARN polymérase se lie pour initier la transcription, se situe avant ou « en amont » du site de démarrage de la transcription. Entre le promoteur et le site de départ de la transcription se trouve la région de l’opérateur.
L’opérateur trp contient le code ADN auquel la protéine répressive trp peut se lier. Cependant, le répresseur seul ne peut pas se lier à l’opérateur. Lorsque le tryptophane est présent dans la cellule, deux molécules de tryptophane se lient au répresseur trp, ce qui modifie la forme de la protéine répresseur pour qu’elle puisse se lier à l’opérateur trp. La liaison du complexe tryptophane-répresseur à l’opérateur empêche physiquement l’ARN polymérase de se lier au promoteur et de transcrire les gènes en aval.
Lorsque le tryptophane n’est pas présent dans la cellule, le répresseur ne se lie pas à l’opérateur, la polymérase peut transcrire les gènes des enzymes et le tryptophane est synthétisé. Comme la protéine répressive se lie activement à l’opérateur pour maintenir les gènes éteints, on dit que l’opéron trp est régulé négativement et que les protéines qui se lient à l’opérateur pour réduire au silence l’expression de trp sont des régulateurs négatifs.
Protéine activatrice du catabolisme (CAP) : Un activateur transcriptionnel
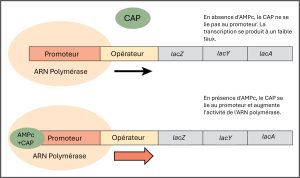
Tout comme l’opéron trp est régulé négativement par les molécules de tryptophane, il existe des protéines qui se lient aux séquences promotrices et qui agissent comme des régulateurs positifs pour activer les gènes. Par exemple, lorsque le glucose est rare, les bactéries E. coli peuvent se tourner vers d’autres sources de sucre pour obtenir du carburant. Pour ce faire, il faut transcrire de nouveaux gènes pour traiter ces sucres alternatifs. Lorsque le taux de glucose baisse, l’AMP cyclique (AMPc) commence à s’accumuler dans la cellule. L’AMPc est une molécule de signalisation qui intervient dans le métabolisme du glucose et de l’énergie chez E. coli. L’AMPc accumulée se lie au régulateur positif, protéine activatrice du catabolisme (CAP), une protéine qui se lie aux promoteurs des opérons qui contrôlent le traitement des sucres alternatifs. Lorsque l’AMPc se lie à la protéine activatrice du catabolisme, le complexe se lie alors à la région promotrice des gènes nécessaires à l’utilisation des autres sources de sucre (figure 16.5). Dans ces opérons, un site de liaison CAP est situé en amont du site de liaison de l’ARN-polymérase dans le promoteur. La liaison CAP stabilise la liaison de l’ARN polymérase à la région promotrice et augmente la transcription des gènes codant pour les protéines associées.
L’opéron lac: Un opéron inductible
Le troisième type de régulation génique dans les cellules procaryotes s’effectue par l’intermédiaire d’opérons inductibles, qui comportent des protéines qui se lient pour activer ou réprimer la transcription en fonction de l’environnement local et des besoins de la cellule. L’opéron lac est un opéron inductible typique. Comme mentionné précédemment, E. coli est capable d’utiliser d’autres sucres comme sources d’énergie lorsque les concentrations de glucose sont faibles. Le lactose est l’une de ces sources de sucre. L’opéron lac code les gènes nécessaires à l’acquisition et à la transformation du lactose provenant de l’environnement local. Le gène Z de l’opéron lac code pour la bêta-galactosidase, qui décompose le lactose en glucose et galactose.
Cependant, pour que l’opéron lac soit activé, deux conditions doivent être remplies. Tout d’abord, le taux de glucose doit être très bas ou inexistant. Deuxièmement, le lactose doit être présent. Ce n’est qu’en l’absence de glucose et en présence de lactose que l’opéron lac sera transcrit (figure 16.6). En l’absence de glucose, la liaison de la protéine CAP rend la transcription de l’opéron lac plus efficace. En présence de lactose, son métabolite, l’allolactose, se lie au répresseur lac et modifie sa conformation de manière à ce qu’il ne puisse pas se lier à l’opérateur lac pour empêcher la transcription. Cette combinaison de conditions est logique pour la cellule, car il serait énergétiquement inutile de synthétiser les enzymes nécessaires à la transformation du lactose si le glucose était abondant ou si le lactose n’était pas disponible. Il convient de mentionner que l’opéron lac est transcrit à un taux très faible même en présence de glucose et en l’absence de lactose.
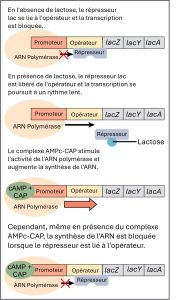
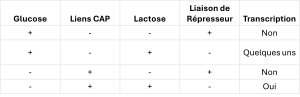
En présence de glucose, la protéine CAP ne parvient pas à se lier à la séquence promotrice pour activer la transcription. En l’absence de lactose, le répresseur se lie à l’opérateur pour empêcher la transcription. Si l’une de ces conditions est remplie, la transcription reste désactivée. Ce n’est qu’en l’absence de glucose et en présence de lactose que l’opéron lac est transcrit (tableau 16.2).
Tableau 16.1. Signaux qui Induisent ou Répriment la Transcription de l’Opéron lac