15.3 Transcription eucaryote
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Énumérer les étapes de la transcription eucaryote
- Discuter du rôle des ARN polymérases dans la transcription
- Comparer et opposer les trois ARN polymérases
- Expliquer l’importance des facteurs de transcription
Les procaryotes et les eucaryotes effectuent fondamentalement le même processus de transcription, à quelques différences près. La différence la plus importante entre la transcription des procaryotes et celle des eucaryotes est due au fait que le noyau et les organites de ces derniers sont liés à une membrane. Les gènes étant dans le noyau, la cellule eucaryote doit pouvoir transporter son ARNm vers le cytoplasme et doit protéger son ARNm de la dégradation avant qu’il ne soit traduit. Les eucaryotes utilisent également trois polymérases différentes qui transcrivent chacune un sous-ensemble différent de gènes. Les ARNm des eucaryotes sont généralement monogéniques, c’est-à-dire qu’ils spécifient une seule protéine.
Initiation de la transcription chez les eucaryotes
Contrairement à la polymérase procaryote qui peut se lier seule à une matrice d’ADN, les eucaryotes ont besoin de plusieurs autres protéines, appelées facteurs de transcription généraux, pour se lier d’abord à la région promotrice et ensuite pour aider à recruter la polymérase appropriée.
Les trois ARN polymérases eucaryotes
Les caractéristiques de la synthèse de l’ARNm des eucaryotes sont nettement plus complexes que celles des procaryotes. Au lieu d’une polymérase unique composée de cinq sous-unités, les eucaryotes possèdent trois polymérases composées chacune de 10 sous-unités ou plus. Chaque polymérase eucaryote nécessite également un ensemble distinct de facteurs de transcription généraux pour l’amener à la matrice d’ADN.
L’ARN polymérase I est situé dans le nucléole, une sous-structure nucléaire spécialisée dans laquelle l’ARN ribosomique (ARNr) est transcrit, traité et assemblé en ribosomes (tableau 15.1). Les molécules d’ARNr sont considérées comme des ARN structurels, car elles ont un rôle cellulaire mais ne sont pas traduites en protéines. Les ARNr sont des composants du ribosome et sont essentiels au processus de traduction. L’ARN polymérase I synthétise tous les ARNr à partir de l’ensemble des gènes ribosomiques 18S, 5,8S et 28S dupliqués en tandem. (La désignation « S » s’applique aux unités « Svedberg », une valeur non additive qui caractérise la vitesse à laquelle une particule se sédimente pendant la centrifugation).
Tableau 15.1. Localisation, produits et sensibilité des trois ARN polymérases eucaryotes

L’ARN polymérase II est situé dans le noyau et synthétise tous les pré-ARNm nucléaires codant pour des protéines. Les pré-ARNm eucaryotes subissent des modifications importantes après la transcription mais avant la traduction. Par souci de clarté, la discussion dans ce module sur la transcription et la traduction chez les eucaryotes utilisera le terme « ARNm » pour décrire uniquement les molécules matures et traitées qui sont prêtes à être traduites. L’ARN polymérase II est responsable de la transcription de la grande majorité des gènes eucaryotes.
L’ARN polymérase III est également situé dans le noyau. Cette polymérase transcrit une variété d’ARN structurels, dont le pré-ARNr 5S, les pré-ARN de transfert (pré-ARNt) et les petits pré-ARN nucléaires. Les ARNt jouent un rôle essentiel dans la traduction ; ils servent de « molécules adaptatrices » entre l’ARNm et la chaîne polypeptidique en croissance. Les petits ARN nucléaires ont diverses fonctions, notamment l’« épissage » des pré-ARNm et la régulation des facteurs de transcription généraux.
Un scientifique qui caractérise un nouveau gène peut déterminer quelle polymérase le transcrit en testant si le gène s’exprime en présence d’α-amanitine, une toxine oligopeptidique produite par l’amanite tue-mouches et d’autres espèces d’Amanita. Il est intéressant de noter que l’α-amanitine affecte les trois polymérases de manière très différente (tableau 15.1). L’ARN polymérase I est totalement insensible à l’α-amanitine, ce qui signifie que la polymérase peut transcrire l’ADN in vitro en présence de ce poison. L’ARN polymérase III est modérément sensible à la toxine. En revanche, l’ARN polymérase II est extrêmement sensible à l’α-amanitine. La toxine empêche l’enzyme de progresser le long de l’ADN et inhibe ainsi la transcription. La connaissance de la polymérase transcriptrice peut fournir des indices sur la fonction générale du gène étudié. Comme l’ARN polymérase II transcrit la grande majorité des gènes, nous nous concentrerons sur cette polymérase dans nos discussions ultérieures sur les facteurs de transcription et les promoteurs des eucaryotes.
Promoteurs et facteurs de transcription de l’ARN polymérase II
Les promoteurs eucaryotes sont beaucoup plus grands et plus complexes que les promoteurs procaryotes. Cependant, tous deux ont une séquence similaire à la séquence -10 des procaryotes. Chez les eucaryotes, cette séquence est appelée boîte TATA et présente la séquence consensus TATAAA sur le brin codant. Il est situé entre -25 et -35 bases par rapport au site d’initiation (+1) (figure 15.10). Cette séquence n’est pas identique à la boîte -10 d’E. coli, mais elle conserve l’élément riche en A-T. La thermostabilité des liaisons A-T est faible, ce qui permet à la matrice d’ADN de se dérouler localement en vue de la transcription. Tous les gènes n’ont pas nécessairement une boîte TATA dans leur promoteur, d’autres éléments de recrutement des facteurs de transcription généraux et de l’ARN polymérase II existent.
Au lieu du simple facteur σ qui aide à lier l’ARN polymérase procaryote à son promoteur, les eucaryotes assemblent un complexe de facteurs de transcription nécessaires pour recruter l’ARN polymérase II sur un gène codant pour une protéine. Les facteurs de transcription qui se lient au promoteur sont appelés facteurs de transcription généraux. Ces facteurs généraux sont tous appelés TFII (pour facteur transcription/polymérase II) avec une lettre supplémentaire (A-J). Le complexe central est le TFIID, qui comprend une protéine de liaison avec la boîte TATA (TBP). Les autres facteurs de transcription généraux se mettent systématiquement en place sur la matrice d’ADN, chacun d’entre eux stabilisant davantage le complexe de préinitiation et contribuant au recrutement de l’ARN polymérase II.
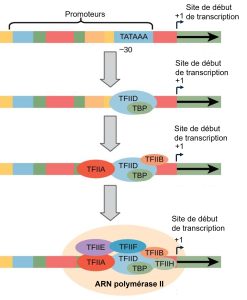
Certains promoteurs eucaryotes possèdent également une boîte CAAT conservée (GGCCAATCT) à environ -80. En amont de la boîte TATA, les promoteurs eucaryotes peuvent également contenir une ou plusieurs boîtes riches en GC (GGCG) ou des boîtes octamères (ATTTGCAT). Ces éléments lient des facteurs cellulaires qui augmentent l’efficacité de l’initiation de la transcription et sont souvent identifiés dans les gènes les plus « actifs » qui sont constamment exprimés par la cellule.
Les facteurs de transcription généraux sont essentiels à la formation d’un complexe de préinitiation sur la matrice d’ADN qui recrute ensuite l’ARN polymérase II pour l’initiation de la transcription. La complexité de la transcription des eucaryotes ne s’arrête pas aux polymérases, aux facteurs de transcription généraux et aux promoteurs. Une armée d’autres facteurs de transcription spécifiques, qui se lient aux séquences d’ADN que l’on appelle enhanceurs et silenceurs (dérivées des expressions “enhancers” et “silencers” en anglais), contribue également à réguler la fréquence à laquelle le pré-ARNm est synthétisé à partir d’un gène. Les enhanceurs et les silenceurs affectent l’efficacité de la transcription, mais ne sont pas toujours nécessaires à son déroulement.
Structures des promoteurs des ARN polymérases I et III
Les processus qui amènent les ARN polymérases I et III à la matrice d’ADN impliquent des ensembles de facteurs de transcription légèrement moins complexes, mais le thème général est le même.
Les éléments promoteurs conservés des gènes transcrits par les polymérases I et III diffèrent de ceux transcrits par l’ARN polymérase II. L’ARN polymérase I transcrit les gènes qui ont deux séquences promotrices riches en GC dans la région -45 à +20. Ces séquences suffisent à elles seules à déclencher l’initiation de la transcription, mais les promoteurs comportant des séquences supplémentaires dans la région comprise entre -180 et -105 en amont du site d’initiation renforceront encore l’initiation. Les gènes qui sont transcrits par l’ARN polymérase III ont des promoteurs en amont ou des promoteurs qui se trouvent à l’intérieur des gènes eux-mêmes.
La transcription eucaryote est un processus étroitement régulé qui nécessite l’interaction de diverses protéines entre elles et avec le brin d’ADN. Bien que le processus de transcription chez les eucaryotes implique un investissement métabolique plus important que chez les procaryotes, il garantit que la cellule transcrit précisément les pré-ARNm dont elle a besoin pour la synthèse des protéines.
LIEN AVEC L’ÉVOLUTION
L’évolution des promoteurs
L’évolution des gènes est un concept familier. Des mutations peuvent se produire dans les gènes au cours de la réplication de l’ADN, et le résultat peut être bénéfique ou non pour la cellule. En modifiant une enzyme, une protéine structurelle ou un autre facteur, le processus de mutation peut transformer des fonctions ou des caractéristiques physiques. Cependant, les promoteurs eucaryotes et d’autres séquences régulatrices de gènes peuvent également évoluer. Prenons l’exemple d’un gène qui, au fil des générations, devient de plus en plus précieux pour la cellule. Le gène code peut-être pour une protéine structurelle que la cellule doit synthétiser en abondance pour une certaine fonction. Si tel est le cas, il serait bénéfique pour la cellule que le promoteur de ce gène recrute plus efficacement les facteurs de transcription et augmente l’expression du gène.
Les scientifiques qui étudient l’évolution des séquences promotrices ont obtenu des résultats variables. Cela s’explique en partie par le fait qu’il est difficile de déterminer exactement où commence et où se termine un promoteur eucaryote. Certains promoteurs se trouvent à l’intérieur des gènes ; d’autres sont situés très en amont, voire en aval, des gènes qu’ils régulent. Cependant, lorsque les chercheurs ont limité leur examen aux séquences du promoteur central humain qui ont été définies expérimentalement comme des séquences qui lient le complexe de préinitiation, ils ont constaté que les promoteurs évoluent encore plus rapidement que les gènes codant pour des protéines.
On ne sait toujours pas comment l’évolution des promoteurs pourrait correspondre à l’évolution de l’humain ou d’autres organismes complexes. Cependant, l’évolution d’un promoteur pour produire plus ou moins d’un produit génétique donné est une alternative intrigante à l’évolution des gènes eux-mêmes.1
Élongation et terminaison chez les eucaryotes
Après la formation du complexe de préinitiation, la polymérase est libérée des autres facteurs de transcription et l’élongation peut se dérouler comme chez les procaryotes, la polymérase synthétisant le pré-ARNm dans le sens 5′ vers 3′. Comme nous l’avons vu précédemment, l’ARN polymérase II transcrit la majeure partie des gènes eucaryotes. Dans cette section, nous nous concentrerons donc sur la manière dont cette polymérase réalise l’élongation et la terminaison.
Bien que le processus enzymatique de l’élongation soit essentiellement le même chez les eucaryotes et les procaryotes, la matrice d’ADN est considérablement plus complexe. Lorsque les cellules eucaryotes ne se divisent pas, leurs gènes existent sous la forme d’une masse diffuse d’ADN et de protéines appelée chromatine. L’ADN est étroitement empaqueté autour de protéines histones chargées à intervalles répétés. Ces complexes ADN-histones, appelés collectivement nucléosomes, sont régulièrement espacés et comprennent 146 nucléotides d’ADN enroulés autour de huit histones comme un fil autour d’une bobine.
Pour que la synthèse des polynucléotides puisse avoir lieu, la machinerie de transcription doit déplacer les histones chaque fois qu’elle rencontre un nucléosome. Cette tâche est accomplie par un complexe protéique spécial appelé FACT, qui signifie « facilite la transcription de la chromatine ». Ce complexe éloigne les histones de la matrice d’ADN au fur et à mesure que la polymérase se déplace le long de celle-ci. Une fois le pré-ARNm synthétisé, le complexe FACT remplace les histones pour recréer les nucléosomes.
La fin de la transcription est différente pour les différentes polymérases. Contrairement aux procaryotes, l’élongation par l’ARN polymérase II chez les eucaryotes a lieu 1 000 à 2 000 nucléotides au-delà de la fin du gène transcrit. Cette queue du pré-ARNm est ensuite éliminée par clivage lors de la transformation de l’ARNm. En revanche, les ARN polymérases I et III ont besoin de signaux de terminaison. Les gènes transcrits par l’ARN polymérase I contiennent une séquence spécifique de 18 nucléotides qui est reconnue par une protéine de terminaison. Le processus de terminaison de l’ARN polymérase III implique une épingle à cheveux de l’ARNm similaire à la terminaison de la transcription indépendante de Rho chez les procaryotes.
Notes de bas de page
1H Liang et coll., « Fast evolution of core promoters in primate genomes »,Molecular Biology and Evolution 25 (2008): 1239–44.
