14.3 Principes de base de la réplication de l’ADN
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Expliquer comment la structure de l’ADN révèle le processus de réplication
- Décrire les expériences de Meselson et Stahl
L’élucidation de la structure de la double hélice a fourni un indice sur la façon dont l’ADN se divise et fait des copies de lui-même. Dans leur article de 1953, Watson et Crick ont écrit un euphémisme incroyable : « Il ne nous a pas échappé que l’appariement spécifique que nous avons postulé suggère immédiatement un mécanisme de copie possible pour le matériel génétique. » Avec des paires de bases spécifiques, la séquence d’un brin d’ADN peut être prédite à partir de son complément. Le modèle à double hélice suggère que les deux brins de la double hélice se séparent pendant la réplication, et que chaque brin sert de modèle à partir duquel le nouveau brin complémentaire est copié. Ce qui n’était pas clair, c’était le mécanisme par lequel les brins se séparaient. Trois modèles ont été suggérés (figure 14.12) : conservateur, semi-conservateur et dispersif.
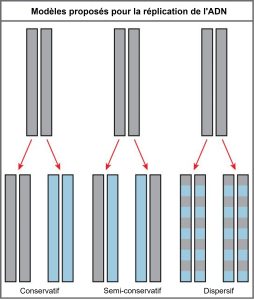
Dans la réplication conservatrice, l’ADN parental reste ensemble et les brins filles nouvellement formés sont aussi ensemble. La méthode semi-conservatrice suggère que chacun des deux brins d’ADN parentaux agit comme un modèle pour la synthèse d’un nouvel ADN ; après réplication, chaque ADN double brin comprend un brin parental ou « ancien » et un « nouveau » brin. Dans le modèle de réplication dispersive, les deux copies d’ADN ont des segments double brin d’ADN parental et des segments double brin d’ADN nouvellement synthétisé intercalés.
Meselson et Stahl étaient intéressés à comprendre comment l’ADN se réplique. Le principe chimique derrière leur expérience était le suivant : un double brin d’ADN ayant incorporé le 15N sera plus lourd, l’ADN ayant incorporé le 14N sera plus léger, et un double brin d’ADN ayant incorporé les deux (14N et 15N) aura un poids moyen en comparaison. Ils ont cultivé la bactérie E. coli pendant plusieurs générations dans un milieu contenant un isotope « lourd » de l’azote (15N), qui est incorporé dans les bases azotées, puis dans l’ADN (Figure 14.13). Donc à ce stade, l’ADN de toutes les bactéries était entièrement composé d’ADN lourd (15N).
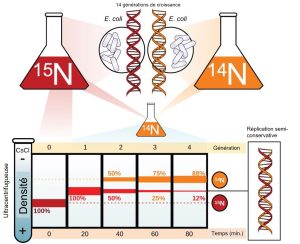
La culture d’E. coli a ensuite été placée dans un milieu contenant du 14N et laissée croître pendant plusieurs générations. Après chacune des générations, un échantillon de bactéries fut récolté et l’ADN fut isolé, puis centrifugé à grande vitesse dans une ultracentrifugeuse. Pour bien séparer l’ADN lourd de l’ADN léger pendant la centrifugation, l’ADN est chargé sur un gradient de sel comme le chlorure de césium ou le saccharose et tourné à des vitesses élevées de 50 000 à 60 000 tr/min. Dans ces circonstances, l’ADN forme une bande en fonction de sa densité flottante : la densité à l’intérieur du gradient auquel il flotte. L’ADN cultivé dans le 15N formera une bande à une position de densité plus élevée (c’est-à-dire plus bas dans le tube de centrifugation) que celui cultivé dans le 14N. Meselson et Stahl ont noté qu’après une génération de croissance dans le 14N après qu’elles aient été déplacées du 15N, la bande unique observée était en position intermédiaire entre l’ADN des cellules cultivées exclusivement dans le 15N et le 14N. Cela suggère un mode de réplication semi-conservatrice ou dispersive. L’ADN récolté sur des cellules cultivées pendant deux générations en 14N formait deux bandes : une bande d’ADN était en position intermédiaire entre le 15N et le 14N, et l’autre correspondait à la bande d’ADN 14N. Ces résultats ne pouvaient s’expliquer que si l’ADN se réplique de manière semi-conservatrice. Et pour cette raison, les deux autres modèles ont donc été exclus.
Lors de la réplication de l’ADN, chacun des deux brins qui composent la double hélice sert de matrice à partir de laquelle de nouveaux brins sont copiés. Les nouveaux brins seront complémentaires aux brins parentaux ou « anciens ». Lorsque deux copies d’ADN filles sont formées, elles ont la même séquence et sont réparties de manière égale dans les deux cellules filles.
