16.6 Régulation traductionnelle et post-traductionnelle des gènes eucaryotes
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Comprendre le processus de traduction et discuter de ses facteurs clés
- Décrire comment le complexe d’initiation contrôle la traduction
- Expliquer les différents modes de contrôle post-traductionnel de l’expression génique
Après avoir été transporté dans le cytoplasme, l’ARNm est traduit en protéines. Le contrôle de ce processus dépend en grande partie de la molécule d’ARNm. Comme nous l’avons vu précédemment, la stabilité de l’ARNm aura un impact important sur sa traduction en protéine. Lorsque la stabilité change, la durée pendant laquelle l’ARNm est disponible pour la traduction change également.
Le complexe d’initiation et le taux de traduction
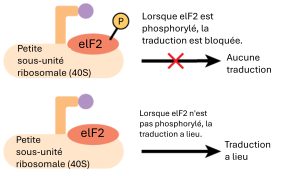
Comme la transcription, la traduction est contrôlée par des protéines qui se lient à l’ARNm et initient le processus. Dans la traduction, le complexe qui s’assemble pour lancer le processus est appelé complexe d’initiation de la traduction. Chez les eucaryotes, la traduction est initiée par la liaison du met-ARNi initiateur au ribosome 40S. Cet ARNt est amené au ribosome 40S par un facteur d’initiation protéique, le facteur d’initiation eucaryote 2 (eIF-2). La protéine eIF-2 se lie à la molécule à haute énergie guanosine triphosphate (GTP). Le complexe ARNt-eIF2-GTP se lie ensuite au ribosome 40S. Un deuxième complexe se forme sur l’ARNm. Plusieurs facteurs d’initiation différents reconnaissent la coiffe 5′ de l’ARNm et les protéines liées à la queue poly-A du même ARNm, de façon à ce que l’ARNm soit en forme de boucle. La protéine eIF4F, qui lie la coiffe, réunit le complexe de l’ARNm et le complexe du ribosome 40S. Le ribosome parcourt ensuite l’ARNm jusqu’à ce qu’il trouve un codon de départ AUG. Lorsque l’anticodon de l’ARNt initiateur et le codon de départ sont alignés, le GTP est hydrolysé, les facteurs d’initiation sont libérés et la grande sous-unité ribosomale 60S se lie pour former le complexe de traduction. La liaison de l’eIF-2 à l’ARN est contrôlée par la phosphorylation. Si l’eIF-2 est phosphorylé, il subit un changement de conformation et ne peut plus se lier à la GTP. Le complexe d’initiation ne peut donc pas se former correctement et la traduction est entravée (figure 16.14). Lorsque l’eIF-2 n’est pas phosphorylé, le complexe d’initiation peut se former normalement et la traduction peut se poursuivre.
Modifications chimiques, activité des protéines et longévité
Les protéines peuvent être modifiées chimiquement par l’ajout de groupements tels que les groupements méthyle, phosphate, acétyle et ubiquitine. L’ajout ou le retrait de ces groupements régule l’activité des protéines ou la durée de leur présence dans la cellule. Parfois, ces modifications peuvent réguler l’emplacement d’une protéine dans la cellule, par exemple dans le noyau, dans le cytoplasme ou attachée à la membrane plasmique.
Des modifications chimiques se produisent en réponse à des stimuli externes tels que le stress, le manque de nutriments, la chaleur ou l’exposition aux rayons ultraviolets. Ces changements peuvent modifier l’accessibilité épigénétique, la transcription, la stabilité de l’ARNm ou la traduction, ce qui entraîne des changements dans l’expression de divers gènes. Il s’agit d’un moyen efficace pour la cellule de modifier rapidement les niveaux de protéines spécifiques en réponse à l’environnement. Les protéines étant impliquées à chaque étape de la régulation des gènes, la phosphorylation d’une protéine (en fonction de la protéine modifiée) peut modifier l’accessibilité au chromosome, peut modifier la transcription (en modifiant la liaison ou la fonction des facteurs de transcription), peut modifier le transport nucléaire (en influençant les modifications du complexe du pore nucléaire), peut modifier la stabilité de l’ARNm (en se liant ou non à l’ARNm pour réguler sa stabilité), peut modifier la traduction (augmentation ou diminution), ou peut modifier les modifications post-traductionnelles (ajouter ou supprimer des phosphates ou d’autres modifications chimiques).
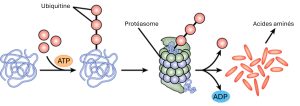
L’ajout d’un groupement d’ubiquitine à une protéine marque la dégradation de cette dernière. L’ubiquitine agit comme un drapeau indiquant que la durée de vie de la protéine est terminée. Ces protéines sont acheminées vers le protéasome, un organite qui a pour fonction d’éliminer les protéines (figure 16.15). L’un des moyens de contrôler l’expression génique consiste donc à modifier la longévité de la protéine.