15.2 Transcription des procaryotes
Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de faire ce qui suit :
- Énumérez les différentes étapes de la transcription des procaryotes
- Discutez du rôle des promoteurs dans la transcription des procaryotes
- Décrivez comment et quand la transcription est terminée
Les procaryotes, qui comprennent les bactéries et les archées, sont pour la plupart des organismes unicellulaires qui, par définition, sont dépourvus de noyau membranaire et d’autres organites. Un chromosome bactérien est un cercle fermé qui, contrairement aux chromosomes eucaryotes, n’est pas organisé autour de protéines histones. La région centrale de la cellule dans laquelle réside l’ADN procaryote est appelée région du nucléoïde. En outre, les procaryotes possèdent souvent d’abondants plasmides, qui sont des molécules d’ADN circulaires plus courtes pouvant ne contenir qu’un ou quelques gènes. Les plasmides peuvent être transférés indépendamment du chromosome bactérien au cours de la division cellulaire et sont souvent porteurs de caractéristiques telles que celles liées à la résistance aux antibiotiques.
La transcription chez les procaryotes (et chez les eucaryotes) nécessite que la double hélice d’ADN se déroule partiellement dans la région de synthèse de l’ARNm. La région de déroulement est appelée bulle de transcription. La transcription se fait toujours à partir du même brin d’ADN pour chaque gène, appelé brin matrice. Le produit de l’ARNm est complémentaire du brin matrice et est presque identique à l’autre brin d’ADN, appelé brin non-matrice. La seule différence entre les nucléotides est que dans l’ARNm, tous les nucléotides T sont remplacés par des nucléotides U (figure 15.7). Dans une double hélice d’ARN, A peut se lier à U par deux liaisons hydrogène, tout comme dans l’appariement A-T dans une double hélice d’ADN.
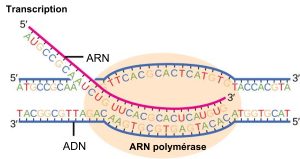
La paire de nucléotides dans la double hélice d’ADN qui correspond au site à partir duquel le premier nucléotide 5′ de l’ARNm est transcrit est appelée le site +1, ou site d’initiation. Les nucléotides précédant le site d’initiation sont désignés par un « – » et sont appelés nucléotides en amont. Inversement, les nucléotides qui suivent le site d’initiation sont numérotés « + » et sont appelés nucléotides en aval.
Initiation de la transcription chez les procaryotes
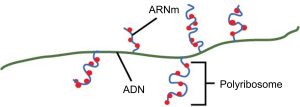
Les procaryotes n’ont pas de noyau fermé par une membrane. Par conséquent, les processus de transcription, de traduction et de dégradation de l’ARNm peuvent tous se produire simultanément. Le niveau intracellulaire d’une protéine bactérienne peut rapidement être amplifié par de multiples événements de transcription et de traduction qui se produisent simultanément sur la même matrice d’ADN. Les génomes procaryotes sont très compacts et les transcrits procaryotes couvrent souvent plus d’un gène ou d’un cistron (une séquence codante pour une seule protéine). Les ARNm polycistroniques sont ensuite traduits pour produire plusieurs types de protéines.
Nous illustrerons la transcription en décrivant ce processus chez Escherichia coli, une espèce bactérienne bien étudiée. Bien qu’il existe certaines différences entre la transcription chez E. coli et la transcription chez les archées, la compréhension de la transcription chez E. coli peut être appliquée à pratiquement toutes les espèces bactériennes.
ARN polymérase procaryote
Les procaryotes utilisent la même ARN polymérase pour transcrire tous leurs gènes. Chez E. coli, la polymérase est composée de cinq sous-unités polypeptidiques, dont deux sont identiques. Quatre de ces sous-unités, appelées α, α, β et β’, constituent l’enzyme centrale de la polymérase. Ces sous-unités s’assemblent à chaque fois qu’un gène est transcrit et se désassemblent une fois la transcription terminée. Chaque sous-unité a un rôle unique ; les deux sous-unités α sont nécessaires pour assembler la polymérase sur l’ADN ; la sous-unité β se lie au ribonucléoside triphosphate qui fera partie de la molécule d’ARNm naissante ; et la sous-unité β’ se lie au brin complémentaire de l’ADN. La cinquième sous-unité, le facteur σ (ou facteur sigma), n’intervient que dans l’initiation de la transcription. Il confère une spécificité transcriptionnelle de façon à ce que la polymérase ne synthétise l’ARNm qu’à partir de site d’initiation spécifique reconnu dans les promoteurs procaryotes. Sans facteur σ, l’enzyme centrale transcrirait à partir de sites aléatoires et produirait des molécules d’ARNm qui spécifieraient un charabia de protéines. La polymérase composée des cinq sous-unités est appelée holoenzyme.
Promoteurs procaryotes
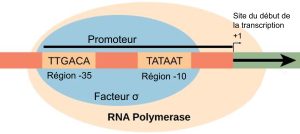
Un promoteur est une séquence d’ADN sur laquelle la machinerie de transcription, y compris l’ARN polymérase, se fixe et initie la transcription. Dans la plupart des cas, les promoteurs se trouvent en amont des gènes qu’ils régulent. La séquence spécifique d’un promoteur est très importante, car elle détermine si le gène correspondant est transcrit tout le temps, une partie du temps ou rarement. Bien que les promoteurs varient d’un génome procaryote à l’autre, quelques éléments sont conservés au cours de l’évolution dans de nombreuses espèces. Dans les régions -10 et -35, en amont du site d’initiation, se trouvent deux séquences consensus du promoteur, c’est-à-dire des régions similaires à tous les promoteurs et à toutes les espèces bactériennes (figure 15.8). La séquence -10, appelée région -10, possède la séquence consensus TATAAT. La séquence -35 comporte la séquence consensus TTGACA. Ces séquences consensus sont reconnues et liées par le facteur σ. Une fois cette interaction établie, les sous-unités de l’enzyme centrale se lient au site. La région -10, riche en A-T, facilite le déroulement de la matrice d’ADN, et plusieurs liaisons phosphodiester sont établies. La phase d’initiation de la transcription se termine par la production de transcrits abortifs, qui sont des polymères d’environ 10 nucléotides qui sont fabriqués et libérés.
Élongation et terminaison chez les procaryotes
La phase d’élongation de la transcription commence par la libération de la sous-unité σ (facteur σ) de la polymérase. La dissociation du facteur σ permet à l’enzyme centrale de se déplacer le long de la matrice d’ADN, synthétisant l’ARNm dans le sens 5′ vers 3′ à une vitesse d’environ 40 nucléotides par seconde. Au cours de l’élongation, l’ADN est continuellement déroulé devant l’enzyme centrale et réenroulé derrière elle. L’appariement des bases entre l’ADN et l’ARN n’est pas suffisamment stable pour maintenir la stabilité des composants de la synthèse de l’ARNm. Au lieu de cela, l’ARN polymérase agit comme un lien stable entre la matrice d’ADN et les brins d’ARN naissants afin de garantir que l’élongation n’est pas interrompue prématurément.
Signaux de terminaison des procaryotes
Une fois qu’un gène est transcrit, la polymérase procaryote doit recevoir l’ordre de se dissocier de la matrice d’ADN et de libérer l’ARNm nouvellement produit. Selon le gène transcrit, il existe deux types de signaux de terminaison. L’un est basé sur les protéines et l’autre sur l’ARN. La terminaison Rho-dépendante est contrôlée par la protéine Rho, qui suit la polymérase sur la chaîne d’ARNm en croissance. Vers la fin du gène, la polymérase rencontre une série de nucléotides G sur la matrice d’ADN et s’arrête. En conséquence, la protéine Rho entre en collision avec la polymérase. L’interaction avec Rho libère l’ARNm de la bulle de transcription.
La terminaison Rho-indépendante est contrôlée par des séquences spécifiques dans le brin complémentaire de l’ADN. Lorsque la polymérase s’approche de la fin du gène en cours de transcription, elle rencontre une région riche en nucléotides C-G. L’ARNm se replie sur lui-même et les nucléotides complémentaires C-G se lient entre eux. Il en résulte une épingle à cheveux stable qui provoque l’arrêt de la polymérase dès qu’elle commence à transcrire une région riche en nucléotides A-T. La région complémentaire U-A de l’ARNm transcrit ne forme qu’une faible interaction avec l’ADN matrice. Ce phénomène, associé au blocage de la polymérase, induit une instabilité suffisante pour que l’enzyme centrale se détache et libère le nouveau transcrit d’ARNm.
Le processus de transcription s’achève à la fin de l’opération. Au moment où la terminaison se produit, la transcription procaryote a déjà été utilisée pour commencer la synthèse de nombreuses copies de la protéine codée, car ces processus peuvent se dérouler simultanément. L’unification de la transcription, de la traduction et même de la dégradation de l’ARNm est possible parce que tous ces processus se déroulent dans la même direction 5′ à 3′ et parce qu’il n’y a pas de compartiment membranaire dans la cellule procaryote (figure 15.9). En revanche, la présence d’un noyau dans les cellules eucaryotes empêche la transcription et la traduction simultanées.