35. Homo neanderthalensis
Homo neanderthalensis (>300 kya)
(“same” / Neander Valley, Germany)

SITES
See “Neandertal sites” map, Figure 35.2
PEOPLE
Too many to mention (see text for some prominent researchers)
INTRODUCTION
One of the best known and most enigmatic of the archaic hominins were the neandertals, Homo neanderthalensis. Although Homo neanderthalensis was originally included in our own genus and species but distinguished by subspecies status, i.e. Homo sapiens neanderthalensis, increasing evidence from DNA analysis suggests that the two lineages split sometime prior to 300 kya and, if new DNA evidence is correct, possibly prior to 800 kya. However, DNA evidence shows that they interbred, possibly as AMH migrated out of Africa one or more times or cohabited with neandertals in the Middle East. Eurasians and Australasians carry, on average, 2.5% neandertal genes. Thus while the RAO model for the origin of AMH is still favored and neandertals are considered a separate species, at least some populations were capable of interbreeding and thus were not true biological species at that point in time and geographic space.
The material that became the holotype for the species was discovered in the Neander Valley near Dusseldorf, Germany. The German word for valley is “thal,” and the “h” is silent. The “h” has been dropped for the common name in some sources. I can only imagine that someone got tired of people pronouncing the “th” and decided to head up a campaign to put a stop to it!
PHYLOGENY
There are two possible scenarios for the origin of neandertals and AMH. The first is based on fossil evidence and the second on DNA. The interpretation of the fossil record suggests that both species are derived from H. heidelbergensis, which in turn likely evolved from a derived form of H. ergaster (possibly H. mauritanicus) in Africa. At some point prior to 500 kya, H. heidelbergensis split into the AMH and neandertal lineages.
The latest genetic evidence supports a split between the AMH and neandertal lineages prior to 800 kya in Africa. The branch leading to neandertals and Denisovans is then considered H. heidelbergensis. The Denisovans and neandertals then split ~640 kya. However, localized groups continued to interbreed.
It is refreshing to learn that populations of hominins have been interbreeding and maintaining or forming genetic relationships since the beginning of “our” time. We modern humans are much more closely related to one another than were those ancient hominin “species” and yet some of us do not see ourselves in others due to physical differences that mean no more than that we went different ways at different times and adapted to different environments.
Regardless of the neandertal/human/Denisovan phylogeny, a group of H. heidelbergensis moved into Western Europe, where a localized group then evolved into the neandertal lineage <300 kya. Transitional forms can be seen in several locales in Western Europe, especially Spain, France, and Germany.
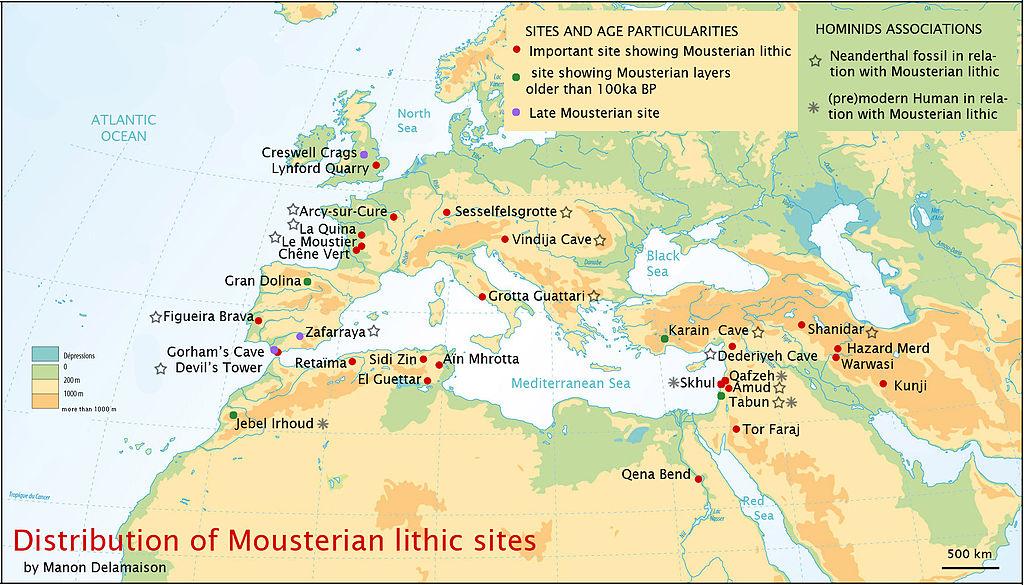
As Pleistocene Europe became colder, neandertals adapted to the harsher conditions. The neandertals from Western Europe, with their stunted and cold-adapted bodies, are known as the “Classic” neandertals, as distinct from those to the east and southeast that retained a more gracile morphology. Dates for the Classic neandertals range from 75 to <30 kya. Figure 35.2 shows neandertal sites in Eurasia.
DISCOVERY AND GEOGRAPHIC RANGE
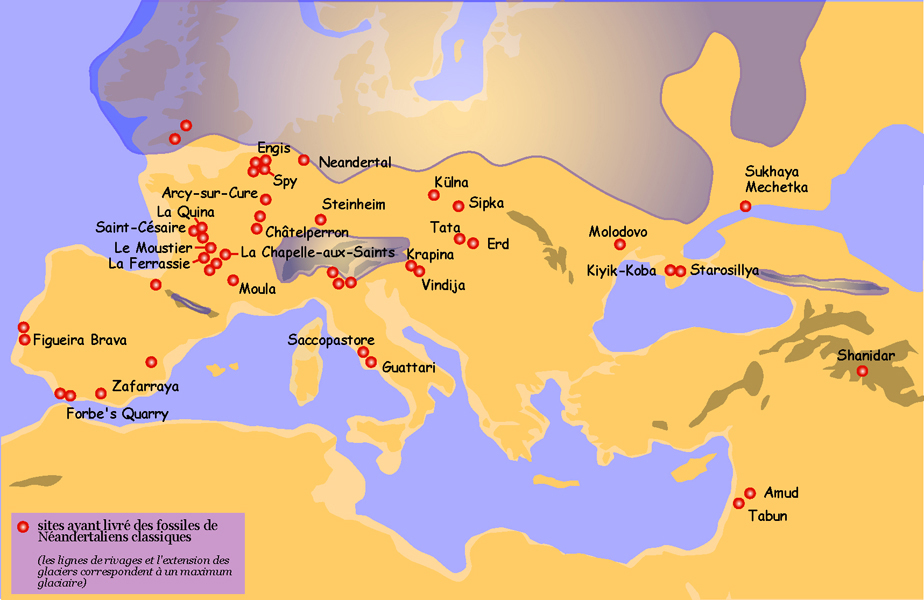
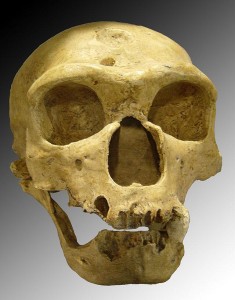
The earliest recognized discoveries were in Belgium and Gibraltar. The next discovery was the Neander Valley remains, which lent the name to the species. Fossil sites are ubiquitous in Western Europe, with the majority located in well-watered river valleys of France. More than 200 sites fall within a 20-mile radius of Les Ezies, France. There are also sites in Germany, Belgium, Spain, Portugal, and Italy. Some of the more famous sites are La Chapelle-aux-Saints, La Ferrassie, and St. Cesaire in France; the aforementioned Neander Valley in Germany; and Zafarraya Cave in Spain. The Chapelle-aux-Saints site has played a key role in the development of the myth of the neandertals as hulking, barbaric cavemen. The remains of an approximately 40-year-old male (see Figure 35.3) were excavated in 1908 and analyzed by Marcellin Boule, who characterized the individual as primitive, brutish, and hunched over. Researchers later realized that the adult was afflicted with arthritis, which accounted for his posture. While we cannot know how neandertals behaved relative to ourselves, they achieved a theretofore unprecedented level of cultural and technological complexity. The derogatory characterization stuck for many years until researchers realized just how much those ancient “peoples” had accomplished, such as intentional burial of their dead.
From their supposed Western European origin, they spread east into the Middle East and as far east as Uzbekistan and northeast to Russia, in the area of the Denisovans. Some researchers do not accept that the nine-year-old boy at the site of Teshik Tash, Uzbekistan is neandertal, but rather they argue that he is AMH.
Non-classic neandertal sites are found in Croatia, the Czech Republic, Hungary, Syria, the Republic of Georgia, Russia, the Ukraine, Iraq, Uzbekistan, and Israel. Famous sites include Krapina and Vindija in Yugoslavia; the cave sites of Kebara, Amud, and Tabun in Israel; Shanidar in Iraq; and the aforementioned Teshik Tash in Uzbekistan.
The Israeli sites have been of interest for decades because they are seemingly contemporary with nearby AMH sites. There has been much speculation as to the nature of interactions between the two species. One theory is that when ice sheets blanketed much of Europe, the neandertals moved down into the Middle East along with other animals. The fact that AMH reached the Middle East by 120 kya but never entered Europe until after 40 kya suggested to some that the neandertals “held” Europe, preventing the encroachment of AMH. When moving down into the Middle East, neandertals may have pushed resident AMH out of the area. During subsequent warmer periods, AMH may have moved back into the area following the retreat of the neandertals to more northerly destinations. This idea of trading places has now been superseded by the idea of contemporaneity and interbreeding, at least by some groups at some point(s) in time.
Figure 35.2 illustrates the broad geographic range of the neandertals. It is likely that during glacial advances, populations moved south so that those in Western Europe were closer to the Mediterranean Sea and eastern neandertals may have pushed down into Israel and other warm areas, along with other animals. The fossil record indicates that animal herds moved up and down in latitude in accordance with climatic pulses, so it is very likely that hominin populations did as well. They were smart and may have inherited past cultural knowledge if they had language and theory of mind and, if for no other reason, they needed to eat and would have followed the game.
By the time that AMH moved into Western Europe ~35 kya, the neandertals had begun to die out. They likely succumbed to the increasingly harsh climate. They also went through an evolutionary bottleneck at some point and lost some of their genetic diversity, possibly leaving them more vulnerable to disease. As in the Middle East, there has been much speculation about what went on when AMH arrived in Western Europe. While they likely carried neandertal genes (unless those western AMH left no modern descendants), they themselves may not have mated with neandertals, and certainly western populations would have appeared somewhat different than Middle Eastern neandertals. However, most eastern neandertals were gone by the time AMH passed through their former eastern geographic range, en route to Western Europe. It has been suggested that AMH outcompeted them either directly, which is known as contest competition, or indirectly, which is known as scramble competition, or possibly even killed them as they encountered them. Contest competition involves one group preventing another group from accessing resources, whereas scramble competition involves one group being better at gaining access to resources than the other. I always think of a bully defending a buffet table from others versus kids scrambling at an Easter egg hunt where some are better than others at getting to and/or finding eggs. It has also been widely accepted that the neandertals were marginalized as AMH encroached on their territory. Except for a more recent date from the Croatian site of Vindija (28 kya), the most recent dates are from the Iberian Peninsula, where it is thought they retreated and died out. Regardless of what transpired between the two species, since it appears that neandertals were on their way out, it is likely a moot point. It is rather fitting that after all of the years of thinking that humans played a role in the demise of the neandertals, it appears they made love not war (at least as far as we know)!
PHYSICAL CHARACTERISTICS
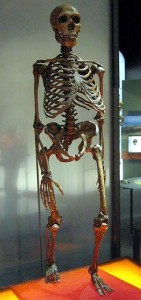
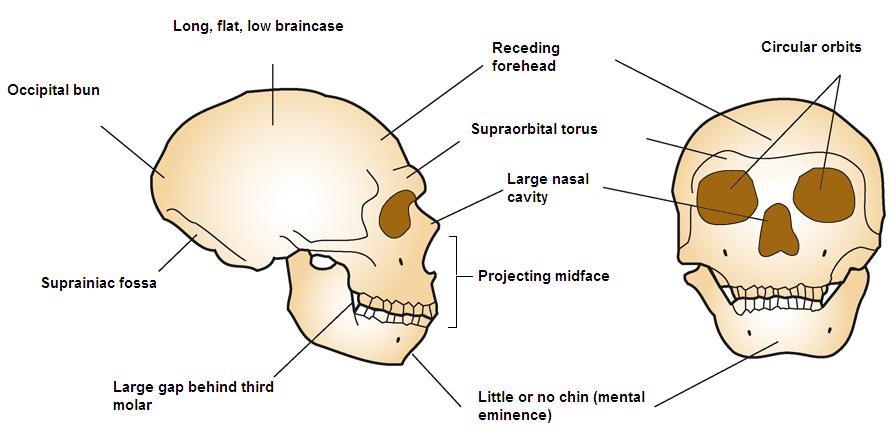
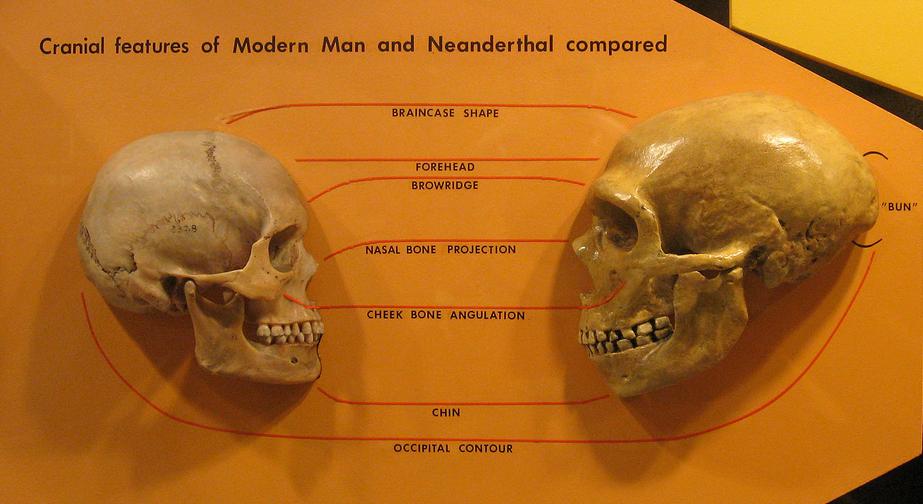
See Figure 35.4 for a full skeletal view of a neandertal. As mentioned, western and eastern neandertals diverged morphologically over time. That clinal variation, i.e. a graded change in physical characteristics over geographic space, is thought to have been the result of time, varying adaptation and exposure to chronically cold Ice Age conditions, and possibly gene flow with AMH in the Middle East. Populations in Western Europe lived at higher latitudes, and the Classic neandertals exhibited cold adaptations that conform to Bergmann’s and Allen’s Rules. Bergmann’s Rule states that as you move away from the equator, mass increases relative to surface area in order to conserve heat, as heat loss is a function of surface area. Allen’s Rule pertains to limb or extremity length, so that organisms in colder environments exhibit shorter appendages. Thus in equatorial Africa, where people have adapted over the long term to hot and dry conditions, body morphology is long and gracile versus the short, stocky morphology of Arctic peoples. In addition to their stocky bodies, short appendages, and barrel chests, neandertals had facial adaptations to the cold. Like H. heidelbergensis, neandertals exhibited midfacial prognathism, large noses, and puffy faces due to enlarged sinuses. Due to their forward-oriented maxilla, the mandible also moved forward, leaving a space behind the third molar, termed a retromolar space. The internal nasal projections were large, thus increasing the internal surface area even further for warming and humidifying inspired air. In addition, unique projections extended from their internal nasal region, up into their orbits. Additional skull characteristics seen in both cold-adapted and eastern neandertals were large, smoothly rolled brow ridges above large, round, widely spaced orbits; “swept back” zygomatics; some unique inner ear characteristics; and, in the occipital region, an occipital bun and suprainiac fossae (two small depressions located above inion, or the external occipital protuberance; see Figure 35.5 for general area). While their skulls were longer and lower than those of AMH (see Figure 35.6), their absolute cranial capacity exceeded even that of modern humans. In accordance with Bergmann’s Rule, a larger brain, while energetically costly in terms of calories, is more conservative from a heat generation and retention perspective. While the neandertal brain was larger, the frontal and parietal lobes (involved with higher thought processes) of AMH were expanded relative to those of neandertals. This may have given AMH an advantage in Ice Age Europe.
Postcranially, neandertals have been described as a cross between a marathon runner (in terms of their endurance) and a wrestler. They were built for chasing down and killing prey. Their upper body was heavily muscled.
ENVIRONMENT AND WAY OF LIFE
The neandertals have traditionally been portrayed as having endured harsh climatic conditions. However, there is debate as to how much of a tolerance they had for conditions in Ice Age Europe. Tattersall (2009) presents an overview of research that suggests that during the coldest periods neandertals lived in more southerly regions and only moved into higher latitudes when temperatures were warmer. While their morphology reflects chronic exposure to cold, like traditional arctic peoples, they did not have the modern technology of those modern humans, such as better shelter, tailored clothes, weapons, and millennia of advanced cultural traditions. They thus had to biologically adapt to the cold. However, when conditions deteriorated as the last glacial maximum approached, they went extinct along with other ill-suited species in the northern latitudes.

It has been known for many years that there were two different types of neandertal settlements, and discussions were often biased as though some inhabited prime real estate and others eked out a living on the open plains. Cave sites in southern France were often described as well-watered river valleys with plentiful caves for shelter, game, water, stone resources for tools, and so forth, and they were thought to have been inhabited for millennia. Open-air sites were apparently inhabited by more nomadic groups that lived out in the open in free-standing structures and followed herd animals. Cave sites and open-air sites are now thought to represent the seasonal shift in subsistence strategies practiced by the same peoples.
Thus while neandertal populations may have been able to live year-round in more temperate regions, the rest were likely semi-nomadic like their ancestors. During the cold of winter in Western Europe, they likely sheltered in southern caves and, in summer, ventured north in pursuit of migrating herds. In addition to caves, they used rock shelters, whereby they built outward from a rock wall or overhang. They did the same inside of caves, building a shelter within a shelter. Post molds form when a wooden post inserted in the ground decays, so that a darker circle of humus is apparent. The pattern of post molds can be used to reconstruct the shape and size of houses and walls in the archaeological record. Based on post molds, neandertals are known to have covered cave entrances, probably during colder periods. Remains of constructed structures show that they used bones, posts, and rocks and likely covered them with skins and insulated with grass. At the Moldova site in the Ukraine, a 26 x 16´ oval ring of mammoth bones was excavated. The bones were likely covered with hides, forming a hut that contained numerous hearths.
During the Pleistocene, European winters are described as having been long and cold and summers were short and cool. Because of the seasonality, plant foods would primarily have been available during warmer months. European neandertals ate a high proportion of meat, with reindeer and mammoth making up the majority of the diet, based upon faunal assemblages and isotopic analyses, respectively. However, dietary composition varied by region. Horses, bovids, and goats inhabited plains whereas at higher elevations, mountain sheep and ibex dominated. At the site of Shanidar, Iraq, faunal remains included goat, sheep, bovid, pig, tortoise, bear, deer, fox, marten, and gerbil bones. At the same site, there is evidence of plant consumption and cooking. Henry (2011) found phytoliths and starch grains in calcium deposits (calculus) on neandertal teeth. Some of the starch from grasses showed damage that is characteristic of cooking. While we know that neandertals used fire, as evidenced by hearths at their sites, and likely ate plants when they were available, it is valuable to finally have supporting evidence. Since Shanidar is south of most of Europe and thus more temperate, it is likely that the neandertals had greater access to such resources.

While debate has raged for some time over whether neandertals practiced cannibalism, fossil material, especially from the French site of Moula-Guercy, provides convincing evidence that at least some groups did eat their own. Neandertal bones at the site exhibit the same signs of processing as animal bones. Bones were disarticulated and hammered open for marrow, and exhibit cut marks from muscle removal.
It is interesting how abhorrent cannibalism is to us. We identify with the neandertals and may feel disappointed that they practiced cannibalism. While it is difficult to say why they ate one another, there is evidence of dietary stress in the form of enamel hypoplasia at some sites, such as Krapina, Croatia. Thus some groups suffered periodic food shortages that resulted in faulty enamel deposition in developing children. If people are starving and there is a dead body available, historic accounts show that they will eat it. Thus it should not be surprising if neandertals consumed the dead, versus killing for consumption. There is no evidence that they practiced cannibalism in all times and places, and thus it could have been in response to extreme conditions.
Prior to the evidence from Moula Guercy, there was great reluctance anytime someone proposed cannibalism to explain damage to remains at particular sites. I think that the stimulus for many of us to feel an affinity for the neandertals is that they intentionally buried their dead. They seem so human. Now that we know that some of our ancestors interbred with them, we may feel an even greater connection to them and will have to accept the good along with possible acts of survival.

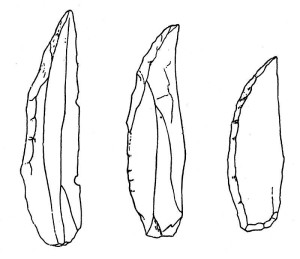
Neandertal culture falls within the period termed the Middle Paleolithic, i.e. the middle portion of the Old Stone Age. The neandertal tool tradition is termed the Mousterian Industry (see Figures 35.9 and 35.10 for examples of Mousterian tools), after the Le Moustier site in France (see Figure 35.8). While there are no known neandertal remains from North Africa, it is of interest that their tools have been found there (see Figure 35.11). The Mousterian method was an improvement on the Levallois technique that provided greater control over resulting flakes. Flakes were then modified into a variety of tools, such as scrapers and points, for various functions. Some of the tools were denticulate, meaning that they were saw-toothed. Like H. heidelbergensis, they made compound tools by hafting stone implements onto handles and shafts.

A later tool tradition (35 kya) from the St. Cesaire site in France is categorized as an Upper Paleolithic industry, as the tools exhibit characteristics of AMH industries. Termed the Chatelperronian tradition (see Figure 35.12), it may be evidence of direct or indirect contact between neandertals and AMH, meaning they obtained the technology via contact with AMH or found one or more tools and used their own methods to replicate them. Some scholars claim the latter versus learning AMH modes of production. Several items from the neandertal site of Arcy-sur-Cure, France, have been interpreted as jewelry, another cultural achievement attributed solely to AMH.
While H. naledi and H. heidelbergensis deposited their dead in deep caves, the neandertals were the first species known to bury their dead in individual graves. Bodies are often found in a flexed position. There is very little evidence of ritual associated with neandertal burials. It appears that they dug a hole, folded the body into the hole, hence the flexed position, and possibly threw some other things in with it. Items are often interpreted as having some significance, but they are usually limited to animal bones and broken tools. However, at the site of Teshik Tash, Uzbekistan, a nine-year-old boy was buried with five sets of wild goat horns that may have adorned his body. While some have suggested that he was an AMH, if he was neandertal it appears to have been a ritualized burial.
The Shanidar site (see Figure 35.13) has always been the most romantic from my perspective. It is a cave site that experienced periodic cave-ins and has yielded the remains of several interesting individuals, some of which were intentionally buried. Shanidar 1 was an adult male. While ultimately the victim of a cave-in, he survived one or more earlier traumatic events in his life. He is thought to have been partially blind due to a head injury that involved one of his eyes. He was missing the end of one of his forearms and thus the hand as well. He suffered a leg injury that resulted in a permanent limp, and some of his teeth were completely worn down. The interesting question is, how did he survive? The oft-cited response is that his group mates helped him in life. He is thus heralded as another case of pre-human altruism or at least kin selection, if the care was provided by his relatives.
Shanidar 3 may be the earliest evidence for murder. Another adult male, he was possibly stabbed, as evidenced by a cut wound to one of his ribs. While the bone showed signs of healing, it is not known whether he died from the wound and was intentionally buried or died in a cave-in. Another interesting burial is Shanidar 4. He was also an adult male that was intentionally buried, and pollen from eight wildflower species was found in the grave. While highly controversial and possibly attributable to a seed-caching rodent species, many would like to think that neandertals not only buried their dead but also placed flowers on their remains.
As I was writing the previous section, it occurred to me that males may have been differentially buried relative to females. While I do not know the answer to that question, it is interesting and food for thought.

The debate as to whether the neandertals could speak has raged for decades. For many years, experts thought that their larynx was situated too high in their throats to have allowed for speech. Our larynx drops during the course of development. Early on it is positioned high in the throat to allow simultaneous drinking and breathing. Babies cannot talk until the larynx drops and they then begin to babble. Thus while they are not mechanically able to speak early on, they are cognitively capable of learning language. Of interest is that some parents now teach their infants sign language so that they can communicate earlier.
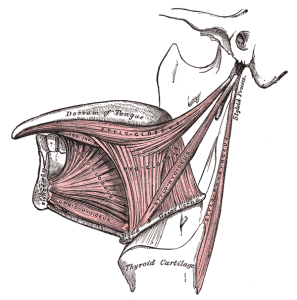
While some researchers still doubt the neandertals’ ability to speak, many have accepted that they likely had spoken language but would have been unable to produce the full range of sounds that characterize our own speech. The discovery of a neandertal hyoid bone at the Kebara site in Israel led many to accept their ability to talk, since its morphology was similar to our own. The hyoid is an important attachment site for the ligaments and cartilages of the larynx and for some extrinsic muscles of the tongue (i.e., geniohyoid, hyoglossus). The most telling evidence in support of neandertal speech, in addition to all of my previous arguments, is the presence of the FOX P2 gene in their genome. We also possess the gene, and it plays an important role in the acquisition of language.

{kind=link}