26. Homo naledi
Homo naledi (date unknown)
(“same” / “star” in Sotho language)
SITE
Rising Star cave system, South Africa
PEOPLE
Cavers Steven Tucker and Rick Hunter and investigated by Paul Dirks and Lee Berger and their associates
INTRODUCTION
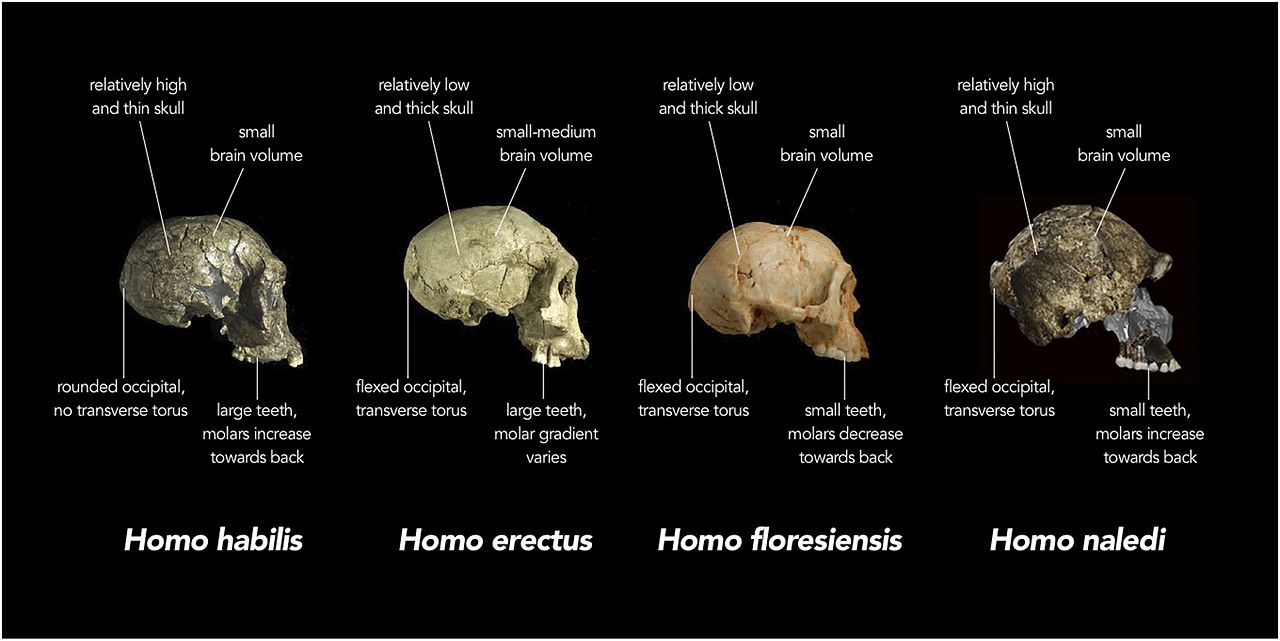
This newest member of our genus has once again confounded the evolutionary history of the Homo lineage. The most exciting aspect is the nature of the remains suggests that they were intentionally deposited in the deep cavern where they were discovered. H. heidelbergensis was heretofore the earliest species thought to have practiced intentional body disposal. Attempts at dating the remains have not been successful. However, Thackeray (2015) has estimated that the species may date to 2.0 ± 0.5 mya, based on comparisons of date and anatomical characteristics among H. naledi, H. habilis, H. rudolfensis, and H. erectus (see Figure 26.2 and Chapter 27 for the erectus grade).
PHYLOGENY
It appears that the majority of researchers agree that the remains reflect a new hominin (see references this section, especially Randolph-Quinney 2015). Like most hominins, the phylogeny of the species is unknown but it likely descended from an australopith ancestry. What makes things even more difficult is that the species shares characteristics with possible extant or near extant species of Homo (H. habilis, H. rudolfensis, and H. erectus), more derived forms (e.g. neandertals and humans), as well as various australopiths. The mosaic of traits is interesting and further supports the bushy nature of the hominin tree.
DISCOVERY AND GEOGRAPHIC RANGE
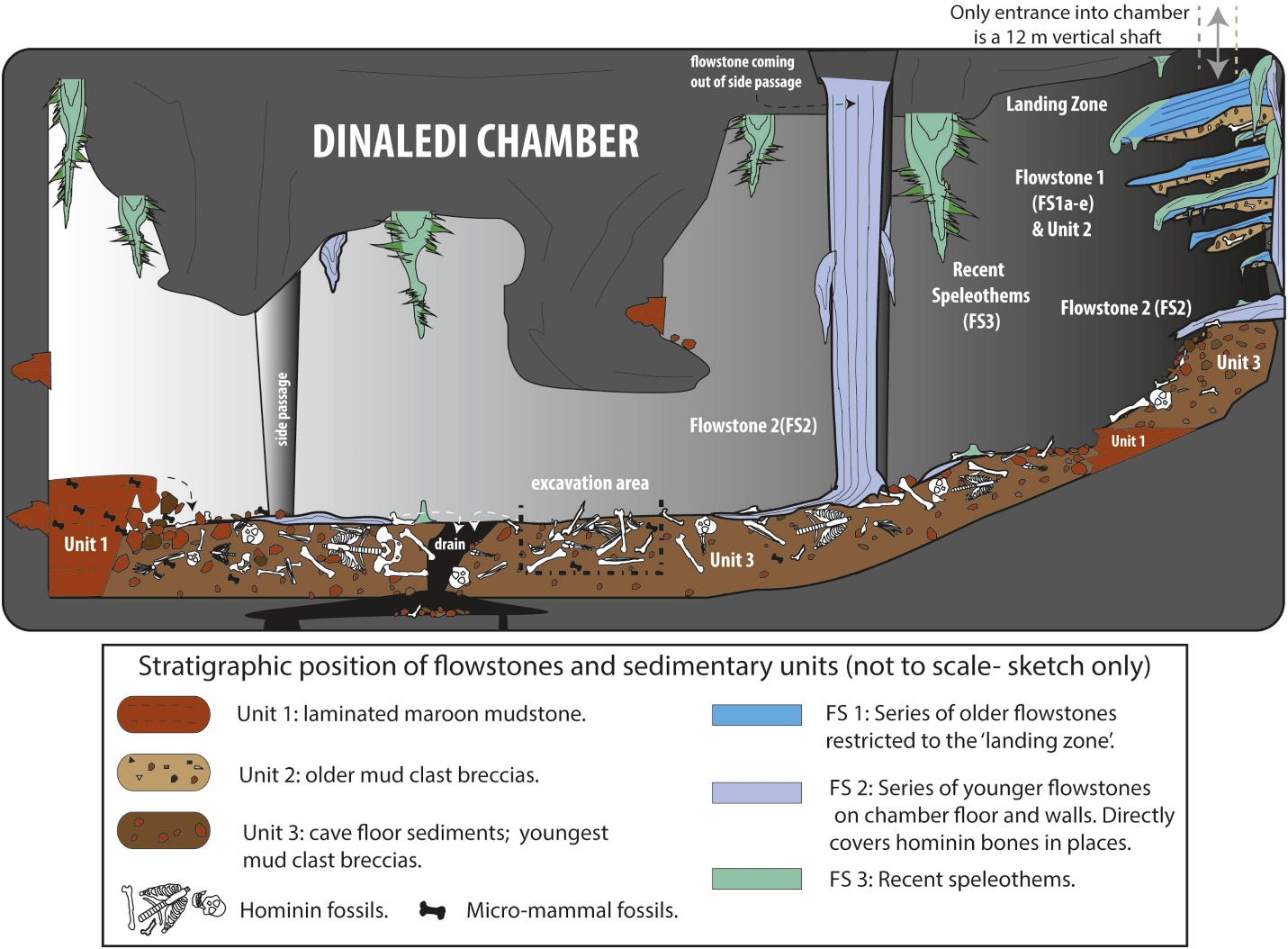
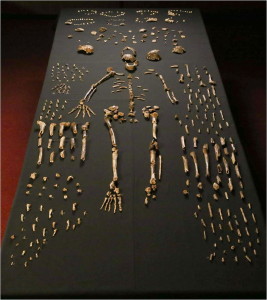
The remains of a minimum of 15 individuals, totaling 1550 fossils (see Figure 26.4), were excavated in 2013 and 2014 from the Dinaledi Chamber, located within the Rising Star cave system in the Cradle of Humankind World Heritage Site, Gauteng Province, South Africa (Berger et al. 2015). The fossils are the largest collection of a hominin species in Africa (Dirks et al. 2015). The chamber is 30 m below ground and is only accessible via a 12 m narrow shaft (Randolph-Quinney 2015 and see Figure 26.3 – top right). Based on depositional data, the bodies were deposited over time (Dirks et al. 2015).
PHYSICAL CHARACTERISTICS
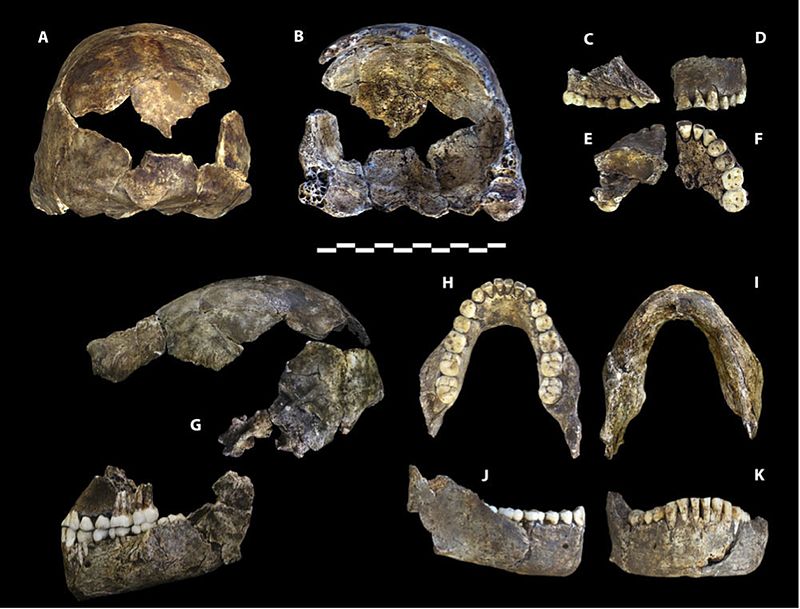
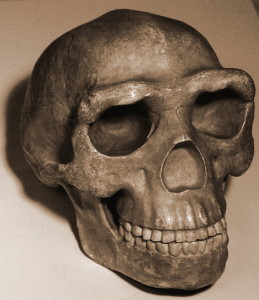
The remains are especially valuable as all body regions are represented, and some bones are articulated, so that anatomical positions and arrangements are preserved, e.g. an almost complete leg of a child and an adult hand (Dirks et al. 2015). The low cranial capacity, elevated shoulder joints, curved phalanges, and trunk and hip morphology are australopith-like. Crania, jaw and teeth morphology, and leg bones are, for the most part, Homo-like. The wrist is most similar to humans and neandertals. The foot is very human-like. (Berger et al. 2015, Harcourt-Smith et al. 2015, Kivell 2015, Thackeray 2015) Thus, we see an able terrestrial biped that could climb, forage, and take refuge in trees.
Cranial capacity falls within the range for the australopiths, with males averaging 560 cc and females, 465 cc (Berger et al. 2015). The base of skull vault is flexed like members of the erectus grade and subsequent species of Homo (see Figure 26.2 and 15.8). The vault bones are thin like those of H. habilis. H. naledi exhibits less postorbital constriction than the earliest australopiths, yet possesses a larger supraorbital torus than any gracile australopith (Berger et al. 2015). Taken together, it is an odd combination. A more gracile face would result in less postorbital constriction, yet the supraorbital torus is associated with chewing stress. Even more surprising, the teeth are smaller than those of H. habilis, H. rudolfensis, and the erectus grade, except for H. floresiensis (Berger et al. 2015). The skull vault is pentagonal in cross-section like Asian H. erectus (see Figure 26.5), due to the presence of a sagittal keel, i.e. a thickening along the midline of the skull, from front to back (Berger et al. 2015). Since the trait is poorly understood in H. erectus, its presence in another species may shed more light on the adaptive significance or ontological processes involved. However, unlike H. erectus, the keel begins in the parietal versus the frontal region (again, see Figure 26.5). While not well developed, H. naledi exhibits a canine fossa, i.e. a depression above the canine tooth, as seen in H. antecessor, H. heidelbergensis, and humans.
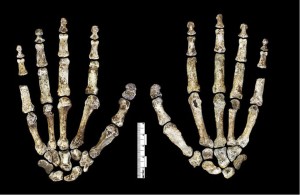
While the hand of H. naledi (see Figure 26.6) shares characteristics with other hominins, the combination of characteristics is unique. They had long fingers and the two more proximal digit phalanges are curved even more than those of australopiths, suggestive of arboreal activities. Yet their wrist morphology is most similar to neandertals and modern humans and, along with their long, robust thumb, they were thus capable of strong manipulatory activities (Berger et al. 2015, Kivell et al. 2015).
The thorax and pelvis were flared like australopiths, but the vertebrae resemble those of the erectus grade and subsequent species of Homo.
While the combination of characteristics seen in the leg bones are distinctive, they are Homo-like, except that the femoral neck is long like that of australopiths. The foot (see Figure 26.7) is very human-like, with the primary differences being the curvature of their digits and less of a medial longitudinal arch (Harcourt-Smith 2015).
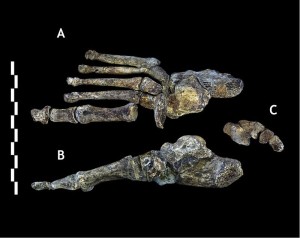
The seemingly advanced bipedal morphology of the two most recent hominin discoveries, i.e. Au. sediba and H. naledi suggest that strong selective forces favored the ability to move through the landscape, in search of food and other resources.
Berger et al. (2015) have calculated the weight of the sexes as follows: males at 55.8 kg and females at 39.7 kg. The only bone that they could use for calculating height was a tibia that yielded an estimate of 144.5 – 147.8 mm. They state that the species falls within the height range of modern small-statured human populations, as well as estimates for the H. georgicus hypodigm from Dmanisi (Lordkipanidze et al. 2007, cited in Berger et al. 2015).
ENVIRONMENT AND WAY OF LIFE
The environment of the region, at around 2 mya, has already been described in the “ENVIRONMENT AND WAY OF LIFE” sections of Au. sediba and P. robustus. Interestingly, the Dinaledi site is 800 m southwest of the P. robustus site of Swartkrans. It was a hominin neighborhood!
The evidence is compelling that the remains could not have been deposited via natural forces, but rather were carried at least part of the way, through a dark and narrow passage. We thus need to reassess our image of the cognitive capabilities and awareness of earlier members of our genus.
