8. Ardipithecus ramidus, Ardipithecus kadabba
GENUS: Ardipithecus (“ground ape”)
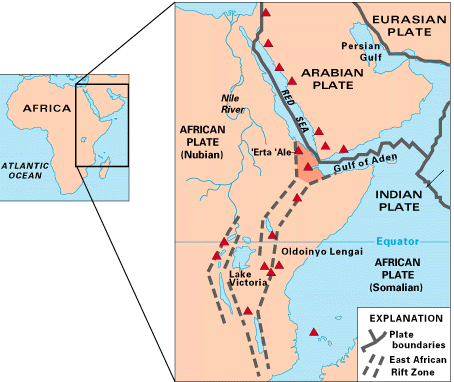
Deposits within the Afar triangle/depression of Ethiopia (see Figure 8.2) have yielded multiple hominin species within the genera Ardipithecus and Australopithecus. This hotbed of hominin fossils is the northern limit of the East African Rift Zone, where the Arabian and African plates converge. The first species of ardipith to be discovered in the area was Ar. ramidus (4.4 mya), and the second and even older species was Ar. kadabba (5.8 mya). When Ar. kadabba was discovered by Yohannes Haile-Selassie, he believed that it was similar enough to Ar. ramidus that he included it in the same genus and species, thus warranting subspecies classification. Since Ar. ramidus was already named, its classification became Ar. ramidus ramidus, in the same manner that we became Homo sapiens sapiens when it was decided that Neandertals should be included in our species and they became Homo sapiens neanderthalensis. The original name is always preserved for the original fossil(s). It becomes known as the “type specimen” or “holotype” and is used to describe the characteristics that define the species. The older subspecies then became Ar. ramidus kadabba. Since that time, they have been split into two species within the genus: Ardipithecus. Each will be discussed below.
Some paleoanthropologists have suggested that Ardipithecus may be a better candidate for our ancestry than one or more of the australopiths. It has also been suggested that australopiths are descended from Ardipithecus or that the ardipiths are a separate but related branch.
Ardipithecus ramidus (4.4 mya)
(“ground ape” / “at the root” in Afar language)
SITES
Aramis, Ethiopia
PEOPLE
Tim White and colleagues
PHYLOGENY
The phylogeny of the ardipiths is unknown. We do not know if they left any descendants, but it is thought that Ardipithecus ramidus is likely descended from Ardipithecus kadabba.
DISCOVERY AND GEOGRAPHIC RANGE
During the early 1990s, fossils were unearthed at the site of Aramis in the Middle Awash region of the Afar Triangle of Ethiopia (see Figure 8.3). Since that time, material from more than 50 individuals has been recovered, in particular the famous “Ardi” skeleton (see Figures 8.4 and 8.5) that is ~50% complete. Prior to the discovery, all or most early African hominin fossils were considered to be australopiths. Tim White and his colleagues determined that the material was distinctive enough to warrant new genus classification.
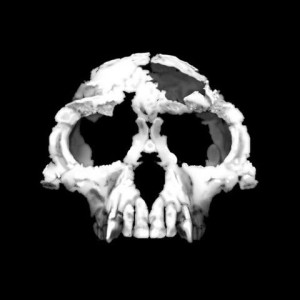
Fifteen years after its discovery, Ardi was presented to the world in a frenzy of media coverage. She shook the world, not just within paleoanthropology, but for anyone interested in our past. She was an upright ape! Yes, we are all apes, but Ardi looked like what we think of as an ape. She had an ape face, small brain, long and strong arms and fingers, and ape-like feet. However, she stood upright with straight rather than flexed legs. The film Discovering Ardi (2009, Discovery Communications) showcases her discovery, fossil processing and analyses, the artist’s skeletal and full-body reconstructions, biomechanical computer-generated graphics, etc.—everything for the modern tech-appreciative student!
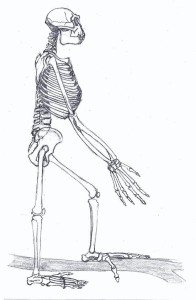
PHYSICAL CHARACTERISTICS
Ardipiths were likely arboreal climbers, like the ancient “basal” proconsulids. However, when they were on the ground, they walked bipedally, albeit with rather clumsy-looking feet. Characteristics that were initially used to designate hominin status were an anteriorly placed foramen magnum, aspects of the innominate and ulna, and especially the morphology of the humerus that suggested that it was not weight-bearing and therefore not involved in terrestrial locomotion.
Primitive characteristics of ardipiths can be seen in many body regions. Their brains were small. Depending on the source, their cranial base (the inferior portion of the occipital bone) was either flat like chimps and gorillas, or angled and tucked under the upper part of the cranium (termed flexed cranial base or basicranial flexion). The enamel on their molars was thin like that of chimps and other extinct forest-dwelling apes. They retained several primitive characteristics that are related to arboreal climbing:
- Elevated shoulder joints for reaching up for branches, etc.
- Long arms and long, curved fingers.
- Mobile wrists.
- Poorly differentiated thumb, meaning that they had poor opposability.
Orangutans are the only great ape with a similar thumb morphology and hence have poor manual dexterity. Ardipiths’ divergent hallux (i.e. the big toe diverges away from the lateral four digits, like our thumbs) would also have been adaptive for climbing.
Derived postcranial characteristics of ardipiths are all in the hip and lower limb. While Ardi’s species had short legs and long arms (i.e. high intermembral index), the morphology of their hip and leg bones reflect bipedality, according to Tim White, Owen Lovejoy, and other members of their team. The innominate and foot morphology combine adaptations for climbing and bipedalism. They were capable of full extension at the knee, unlike the flexed knee seen in extant apes. Their feet were stable and supported their body weight and the divergent hallux facilitated grasping and climbing.
Body mass is estimated from Ardi’s skeleton and thus no estimate for males of the species is possible. Ardi is estimated to have been 3′11” (120 cm) tall and weighed about 110 lb (50 kg) (Gibbons 2009). The absence of both a honing complex and pronounced prognathism suggests to some researchers, such as C. Owen Lovejoy, that males were not competing for females and may have formed pair-bonds with them.
Review of Primitive Characteristics
- Small brain and flat cranial base.
- Thin molar enamel.
- Climbing complex of upper limb:
- Elevated shoulder joint.
- Long arms.
- Long and curved fingers.
- Low manual dexterity.
- Divergent hallux.
Review of Derived Characteristics
- Anteriorly oriented foramen magnum.
- Bipedal hip, leg, and foot characteristics.
ENVIRONMENT AND WAY OF LIFE
The sites where ardipith fossils have been recovered were a mosaic environment consisting of wood- and grasslands during the late Miocene and early Pliocene. Those ancient apes likely subsisted on a combination of arboreal and terrestrial forest resources. Their thin molar enamel suggests that, like chimps and orangutans, their diet consisted of relatively soft food items, such as fruit, young leaves, and shoots and possibly ants and termites. They could locomote within and between trees when foraging, or climb down and travel between trees or forested areas. Since all great apes build nests, ardipiths may have made a new arboreal nest every night.

Some scientists see the lack of canine sexual dimorphism and honing complex as evidence for pair-bonding. There is good evidence from the animal literature to relate sexual monomorphism (no difference in size between males and females) and pair-bonding. As mentioned in Chapter 4, while there are several groups of primates that exhibit that grouping and mating strategy, they are all arboreal. The lesser apes are arboreal, medium-sized (<6 kg) primates that form territorial pairs. Their mode of locomotion is brachiation, i.e. swinging under branches. While that is a more efficient means by which to avoid predation relative to ardipiths climbing around in the trees, the latter were larger and heavier and thus were likely fairly safe. When they descended to the ground, however, pairs would have been fair game for predators.
The idea that male provisioning, in combination with pair-bonding, was the evolutionary stimulus for bipedalism is also problematic. As I discussed in Chapter 5, I do not understand why females would need to be provisioned unless their babies could no longer hang on. I just do not see males risking life and limb to bring food back to females. I know of no examples in nature of a mammalian male provisioning his mate. It is more likely that resources became fragmented to the point that upright locomotion was the most efficient way to move between trees and forest patches. Bipedalism was an adaptation to enable continued reliance on forest resources while minimizing competition. Ardipiths could move to another forest patch when resources became scarce. I think of them as being semi-terrestrial apes like chimps and gorillas but, unlike those extant species, they had less of a stable resource base. They thus evolved a more efficient locomotor strategy for expanding their home range.

Ardipithecus kadabba (5.8 mya)
(“ground ape” / “oldest ancestor” in Afar language)
Yohannes Haile-Selassie discovered the second ardipith species in the Middle Awash region of the Afar Depression (see Figure 8.3). The species is thought to be ancestral to Ar. ramidus. Dental characteristics are more ape-like than those of Ar. ramidus. While the fossils are very fragmentary, a toe bone suggests bipedal foot movement involving the “toe-off” motion, i.e. when we push off with the toes of one foot as we plant our other foot following its “swing” phase.