3 Body Temperature and Thermoregulation

Amphibians and reptiles are neither warm-blooded or cold-blooded
Amphibians and reptiles are often called “cold-blooded” animals. Mammals and birds, in contrast, are often called “warm-blooded”. This dichotomy is somewhat misleading because while it is true that birds and mammals can remain relatively warm regardless of the ambient temperature, it is not true that amphibians and reptiles are systematically colder than their feathery and furry relatives. In fact, a lizard basking in the sun can have a body temperature equal or higher than a mammal. The key difference between birds and mammals, on one hand, and amphibians and reptiles, on the other, is not the temperature of the body (or the blood), but where the heat comes from and how much the temperature of the body varies over time.
Amphibians and reptiles are ectotherms (ecto = outside, and therm = temperature). Ectotherms use external sources of heat like the sun to maintain their body at a desired temperature. In contrast, birds and mammals are endotherms (endo = inside). Endotherms use their cellular machinery to maintain a constant body temperature that is typically well above the ambient temperature. For instance, your core temperature remains at 37.5°C regardless of the ambient temperature.
Animals maintaining a constant body temperature are called homeotherm (homeo = same). The body temperature of amphibians and reptiles tends to fluctuates over time. Animals experiencing fluctuating body temperature are called poikilotherms (poikilo = variable). Note that poikilotherm is not a synonym of ectotherm. Likewise, homeotherm is not a synonym of endotherm. Poikilothermy and homeothermy refer to the fluctuation in body temperature. In contrast, ectothermy and endothermy refer to the dominant source of body heat. Confusion between these terms comes from the fact that most ectotherms are also poikilotherms and most endotherms are also homeotherms but there are many exceptions. For instance, a bat undergoing daily torpor can be considered an ectotherm poikilotherm. Similarly, an aquatic reptile living a tropical area that experiences little temperature fluctuations daily and seasonally is closer to a homeotherm than to a poikilotherm.
Ectothermy has its advantages
Ectothermy influences the ecology and behaviour of amphibians and reptiles directly. For instance, most terrestrial ectotherms cannot migrate over long distances (some insects are exceptions) or be active at cold or sub-zero temperatures like mammals and birds. Ectothermy is the ancestral condition in tetrapods while endothermy is a derived trait that evolved independently in mammals and birds. It may thus be tempting to think of reptiles and amphibians as “primitive” or less adapted than birds and mammals. This is view is incorrect. As we will see, amphibians and reptiles thrive under conditions that are lethal to birds and mammals.
Fuel economy is a key advantage of ectothermy. For example, a lizard can live and reproduce on approximately 10% of the energy that a mouse of the same weight needs. In birds and mammals, most of the energy obtained from food is used to keep the body warm. Endothermy is expensive and it gets more expensive as an animal gets smaller. Small birds and mammals like hummingbirds and shrews have extremely high metabolic rates because they have a high surface area to volume ratio which causes them to rapidly lose their body heat. The metabolic tax for being small is not nearly as steep for ectotherms. The smallest frog species is nearly 200 times lighter that the smallest mammal species.

Performances varies with body temperature
Take a lizard and measure how fast it can run at different temperatures. Then make a graph showing running speed as a function of body temperature. Your graph will look like the one below.

Running speed (a measure of performance) will slowly increase with body temperature until it reaches a plateau after which it will decrease rapidly. The curve is asymmetrical (not bell-shaped). The range of temperatures at which the lizard is running the fastest (the plateau) is the optimal temperature for running speed. If the temperature is too low or too high, the lizard stops running completely and may die. These extremes are called the lethal temperatures.
If you make similar curves for other functions such as digestion speed, or egg development, you will get curves looking similar to the running speed curve. Physiologists have often compared thermal performance curves for multiple bodily functions for the same species. In general, the different functions peak around the same optimal temperature. The different functions are thus said to be co-adapted.

It makes sense that several important functions have similar thermal performance curves. Indeed, it would be challenging for an animal to performance optimally if running speed peaked at 30°C but digestion peaked at 20°C. As we can see from the curves above, slightly exceeding the optimal temperature can have lethal consequences so herps tend to avoid temperatures above their optimum. One exception is to create what is called a behavioural fever. When infected by a pathogen, certain amphibians and reptiles will select higher than optimal temperatures. This behaviour presumably helps fight the infection by forcing the pathogen outside its optimal temperature for growth and reproduction.
Under certain circumstances, it may be advantageous for herps to remain below their optimal temperature to reduce their energy consumption. As the ambient temperature drops, mammals, and birds must spend more energy to maintain their high body temperature. Herps, on the other hand, spend less energy as it gets cooler because their metabolic rate is a direct function of their body temperature. This can be very advantageous because food tends to be scarcer when it is colder (e.g. in winter). Being cool is also advantageous for herps overwintering underwater because it reduces their demand for oxygen (more on this in the overwintering unit).
Thermoregulation
To grow and reproduce successfully, amphibians and reptiles must spend much of their time budget close to their optimum body temperature. If we map the diversity of amphibians and reptiles for the whole world, we can see that that most species live in or near the tropics where temperatures are relatively warm and stable year round. Thus most species of amphibians and reptiles can remain close to their optimal temperature without having to invest much time and energy in behavioural thermoregulation.
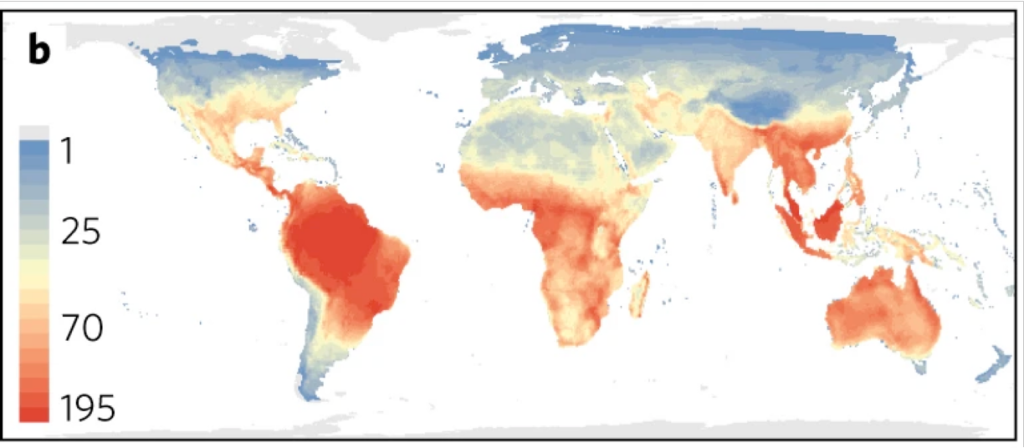
Temperate areas are a different story however. As we move away from the equator to higher latitudes (or to higher altitudes), temperatures become more variable both daily and seasonally. At our latitude, the temperature ranges from lethally cold to lethally hot in a matter of weeks, and for most of the year the ambient temperature is well below the optimal body temperature of most species. In temperate deserts (e.g. The Chihuahua, Sonora, and Great Basin), daily variation in temperature are even more pronounced. Such environments are considered thermally challenging because it is difficult for amphibians and reptiles to reach and remain in their thermal sweet spot. Under such conditions, thermoregulation is essential. Behaviour is the main way by which amphibians and reptiles regulate their body temperatures but some species also use physiological tricks to control the rate at which they warm up or cool down.
Gaining and Loosing Heat
The body temperature of amphibians and reptiles is a direct function of the temperature of the environment, but that does not mean that their body temperature is always the same as the environment. At least in temperate areas, the body temperature of amphibians and reptiles is rarely the same as the ambient temperature. Fully aquatic species are an exception. Body heat is rapidly lost in water (through convection) so the body temperature of fully aquatic ectotherms is generally the same as the temperature of water they are in.
Heat can be gained or lost through different routes. We can summarize the total heat energy gained with the following equation:

Where:
Qabs = Radiation absorbed by the surface of the animal
M = Metabolic heat generated by the body
R = IR radiation received or emitted by the animal surface
C = Heat gained or lost by convection (i.e. through the flow of air or water over the animal)
G = Conduction – heat gained or lost by direct physical contact with objects or water
LE = Heat gained by condensation or lost by evaporation
The units are watts per square meter of animal surface
Amphibians and reptiles use behaviours, and to a lesser extent, physiological tricks to increase or decrease how much heat energy is gained or lost through these routes.
Basking Behaviour
Basking is a quintessentially reptilian behaviour. At high latitudes, activity seasons are short, and the daily temperatures are typically cool so temperate species of amphibians and reptiles tend to frequently bask to extend the amount of time they spend at their optimal temperature. Basking is a more complex behaviour than simply sitting in the sun. It involves various behavioural and physiological adjustments. Let’s look at how some amphibians and reptiles use behaviour and physiology to modulate heat exchange during basking.
In the picture below, a male Collared Lizard (Crotaphytus collaris) is basking under the midday June sun in the hot Sonoran desert (Arizona). Its back is facing the sun, so he is gaining heat directly from the intense solar radiations (Qabs). However, his upright posture limits how much heat is gained. If he was laying flat on the ground, he would be gaining more heat from the sun but by resting upright he is reducing the surface area of its body directly exposed to the sun. The rock he is standing on is also gaining heat from the sun and some of that heat is transferred to him via two routes. Firstly, the rock transfers heat by conduction (G) to to every parts of the lizard’s body in direct contact with it. Secondly, some of the solar radiations hitting the rock are reflected back to the lizard in the form of infra-red radiations (R). If you look closely at the lizard’s hind feet, you will see that his toes are curled up. That is presumably to reduce heat transfer by conduction. The same can be said of every parts of his body not touching the rock. Our lizard’s posture also promotes heat loss. By keeping his body off the rock, he exposes much of it to the wind. As the air flows around his body, heat is lost by convection (C). If this lizard was laying flat on the rock, he would rapidly overheat in this environment but by adjusting his posture, he can maintain its optional temperature for longer which is giving him more time to keep an eye out for females, interlopers (they are territorial) and bugs to eat.

Our collared lizard’s main thermoregulatory concern was avoiding overheating but sometimes the main concern is gaining heat as fast as possible. Let’s move from Arizona to southern Saskatchewan where a female Greater Short-Horned Lizard (Phrynosoma hernardesi) is basking on a cool spring morning. In contrast to the collared lizard, the short-horned lizard lays flat against the ground. This posture allows her to maximize heat gain from solar radiation (Qabs), as well as from direct contact with the ground (G). This flattened posture also allows her to reduce heat loss via convection from the wind (C). Being close to the ground reduces convective heat loss because a layer of still air called the boundary layer forms just above the ground. Every physical object has a boundary layer around it. The thickness of the boundary layer depends on the size of the object as well as on the flow of air around it. The bigger an object, the thicker the boundary layer. The slower the air flow, the thicker the boundary layer. The air in the boundary layer above the ground is warmed up by the heat radiating from the ground and other objects (plants, rocks, logs). The rocks and plants near the surface of the ground also break the wind and thus reduce heat loss by convection in the boundary layer. By staying close to the ground, this female short-horned lizard remains in the relatively warm boundary layer of the ground and minimizes heat loss by convection (C). Short-horned lizards in the mountains of Texas can elevate their body temperature to nearly 30°C when air temperature is as low as 1.5°C by remaining in the boundary layer of the ground. The same is probably true in southern Alberta and Saskatchewan where this species is also found.

Some amphibians and reptiles exploit the cooling effects of evaporation during basking. As water evaporates from a surface, heat is lost in a process called evaporative cooling (LE). Humans, exploit this process to cool down. That’s what sweating is all about. Amphibians and reptiles do not have sweat glands but can nonetheless experience evaporative cooling by exposing the moist surfaces of their body to the air. The skin of amphibians is thin and permeable (a requirement for cutaneous respiration) so amphibians have in theory a high potential to use evaporation to cool down. The problem with evaporative cooling thought is the evaporation bit. Amphibians using evaporation to cool down must replenish the water they loose through evaporation or they risk of becoming critically dehydrated which would affect, among other things, their ability to breath through their skin. Amphibians do not drink so the water lost by evaporation must be re-gained through cutaneous absorption. Amphibians can do this by sitting on a wet surface or in water. Many species of semi-aquatic frogs like the American Bullfrog (Lithobates catesbeiana), the Leopard Frog (Lithobates pipiens), and the Green Frog (Lithobates clamitans) are active during the day. On warm sunny days, these species are often observed sitting on the margin of ponds and lakes with their limbs partly submerged . Such species appear to use evaporative cooling to maintain a stable body temperature under warm conditions but can readily regain the water lost by evaporation.

Reptiles also exploit evaporative cooling but because their skin is largely impermeable, they do so by gaping or panting to expose the permeable lining of their mouth to the air. For instance, gaping allows desert Iguanas (Disposaurus dorsalis), to maintain a body temperature 3°C lower than the ambient temperature, and a head temperature 6°C lower than ambient. Basking turtles are often observed gaping, but it is unclear if this behaviour has a thermoregulatory function (perhaps you will figure it out for your master’s).

Habitat Selection
Animals don’t typically use their environment at random but rather select certain habitat features over others. This process is called habitat selection. Temperature can play an important role in habitat selection by amphibians and reptiles and when and where amphibians and reptiles are found is influenced by the need to thermoregulate. Within the habitats available to a particular individual, temperatures vary both spatially and temporally. For instance, it is warmer at the edge of a field than a few meters in the forest. Similarly, it is warmer on top of a rock than underneath that same rock. .
As we have seen above, amphibians and reptiles have an optimal body temperature. We can thus expect them to favour habitats in which their optimal body temperature can be reached and maintained. This prediction appears to be true in many species living in temperate areas. For instance, the Eastern Milk Snake (Lampropeltis triangulum triangulum) in Ontario uses open habitats more than they are available. These habitats also have the best opportunities to maintain a body temperature close to their optimum temperature.


Temperature selection is also affected by an animal’s reproductive, digestive, and health status. For instance, digesting snakes seek warmer temperature than non-digesting ones. Gravid reptiles, at least in temperate areas, also seek warmer temperature. For instance, northern water snake females in Ontario give birth in August. In July and August, the gravid females maintain a higher body temperature and spend more time near their optimum temperature than males and non-gravid females.


Physiological Mechanisms of Thermoregulation
Amphibians and reptiles have a very limited ability to generate heat but they can nonetheless use physiological processes to adjust the speed at which they gain and loose heat. Thus physiological thermoregulation in amphibians and reptiles largely refers to mechanisms used to buffer heat exchanges with the environment rather than mechanisms used to generate metabolic heat. There are however a few notable exceptions. A few large species of reptiles can generate substantial metabolic heat. We will briefly discuss those exceptions at the end of this section.
Color change

Changing color is a common way by which amphibians and reptiles modulate heat gain. By changing color, animals change the absorptivity of their integument and thus the rate at which they gain heat from solar radiations. As you know, darker surfaces absorb more radiation and thus heat up faster than lighter surfaces. Rapid color change is achieved by the migration of melamine pigments inside pigment bearing cells called chromatophores. When the pigments are concentrated deep in the cell, the animal looks pale. When the pigments are spread all over the cell, the animal looks dark.
Some amphibians and reptiles darken their skin to speed up heat gain and lighten their skin to reduce it. For instance, the bearded dragon (Pogona vitticeps) changes color as they change body temperature. They are dark when cool but become paler as they warm up. Color change affects skin reflectivity by up to 27% and can speed up warming by 22 minutes per active day. Changing color allows them to spend more time at their optimal temperature which presumably increases their fitness.

Cardiovascular Adjustments
Blood carries a lot of important stuff through out the body including oxygen, nutrients, hormones, and metabolic wastes. Blood also moves heat around the body and some reptiles can adjust their blood flow to modulate the rate at which they warm up or cool down. This mechanism of physiological thermoregulation has been well studied in bearded dragons (Pogona vitticeps). When these lizards begin to bask early in the day, the air temperature is relatively cool and solar radiations relatively weak. To warm up faster, bearded dragons dilate their skin capillaries which sends more blood to the skin; a phenomenon called skin perfusion. The skin is directly exposed to the sun so the blood just under its surface warms up as it delivers oxygen to the skin. The warmed blood then returns to the heart via veins where it warms up the core of the animal. Skin perfusion is only part of the story though. The blood returning from the skin is de-oxygenated. This blood would normally go to the lungs via the pulmonary artery but that is not what happens. Instead of going to the lungs, some of the warmed but de-oxygenated blood returning from the skin goes back to the skin without going to the lungs! The blood returning from the skin by-passes the pulmonary artery and re-enters the systematic circulation via the left and right aortas. This rerouting is called a right to left cardiac shunt because the blood arriving to the left atrium is shunted to the right side of the heart instead of remaining on the left side. By-passing the pulmonary circulation reduces blood cooling at the lung interface and speeds up warming.
 |
 |
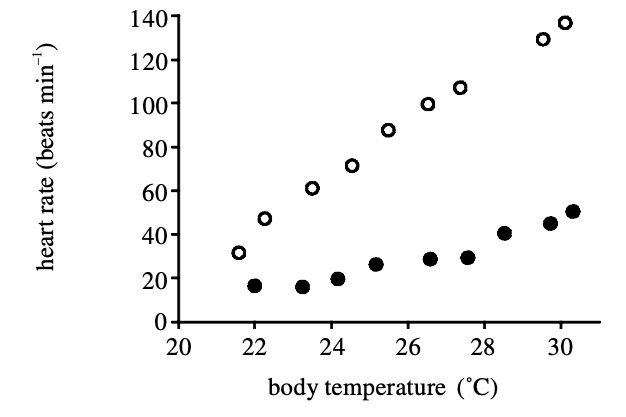
Cardiac shunting is possible in reptiles (lepidosaurs and turtles) because their ventricle is not fully divided in two as in birds and mammals. In mammals, and birds, the de-oxygenated blood entering the right atrium can only go to the right ventricle which sends it to the lungs via the pulmonary artery. In reptiles, the de-oxygenated blood flowing from the right atrium to the right ventricle can “hops” over the incomplete septum between the right and left ventricles and re-enter the systemic circulation without going to the lungs first. Skin perfusion and the right to left cardiac shunt are accompanied by an increase in heart rate. Indeed, at the same temperature, a warming lizard has a faster heart rate than a cooling one; a phenomenon called heart rate hysteresis.

Metabolic Heat
Reptiles and amphibians don’t produce enough metabolic heat to raise their body temperature above the ambient temperature but there are at least two exceptions to this: the Leatherback Sea Turtle and some species of pythons.
The Leatherback Sea Turtle (Dermochelys coriacea) is the largest living species of turtle with adults weighting up to 850 Kg. Their large size gives them a certain thermal inertia meaning that their internal temperature takes a long-time to change when the ambient temperature does. This is very useful for a reptile frequenting the cold waters of the globe. Leatherback sea turtles are regularly observed in Newfoundland and some individuals have been reported in Alaska, Iceland, and Norway. Moreover, Leatherbacks can dive down to 1200 m where the water temperature hovers just above the freezing point.

Being a giant is not the only thing allowing Leatherbacks to keep their body temperature well above the water temperature. Given enough time, any large ectotherm would cool down to the water temperature but Leatherbacks don’t. In the North Atlantic, Leatherbacks can keep their body temperature 12°C above the water temperature. They achieve this feat through both behaviour and physiology. Firstly, they take advantage of the warmer surface water (behaviour) to raise their body temperature. Secondly, the contractions of their powerful fins muscles during swimming as well as their digestion (physiology) generate enough heat to warm up their large bodies. Leatherbacks can thus be considered somewhat endothermic.

Interestingly, Leatherbacks generate so much heat from contacting their fin muscles that they would overheat if that heat was to be brought to the turtle’s core with their venous blood. To prevent this, Leatherbacks cool their blood with a counter current heat exchange system located at the base of their limbs. In the limbs, arteries and veins run parallel to each other. The heat generated by the muscles, and carried by the venous blood, gets transferred to the arteries bringing the oxygenated blood to the muscles. This system keeps the swim muscle warm but cools down the blood in the veins returning to core.

Pythons also use muscle contractions to generate heat, although in this case the snakes are not trying to raise their own body temperature but that of their eggs. Indeed, females from several species of pythons brood their eggs. While brooding, they rapidly contract their muscles which generates enough heat to raise the temperature of their eggs above the ambient temperature. For instance, Burmese pythons (Python bivittatus) can keep their eggs 4°C above ambient. Warming up the eggs has the advantages of speeding up their development and reducing the incubation period but it comes at an important cost. At 25.5°C the metabolism of a brooding mother is about twenty times higher than a non-brooding snake at the same temperature.

References
Brown, G. & Weatherhead, P.J. (2000). Thermal ecology and sexual size dimorphism in northern water wnakes, Nerodia sipedon. Ecological Monographs. 70. 311-330. 10.1890/0012-9615(2000)070[0311:TEASSD]2.0.CO;2.
Casey, J. P., James, M. C., & Williard, A. S. (2014). Behavioral and metabolic contributions to thermoregulation in freely swimming leatherback turtles at high latitudes. Journal of Experimental Biology, 217(13), 2331–2337. https://doi.org/10.1242/jeb.100347
Davenport, J., Jones, T. T., Work, T. M., & Balazs, G. H. (2015). Topsy-turvy: turning the counter-current heat exchange of leatherback turtles upside down. Biology Letters, 11(10), 20150592. https://doi.org/10.1098/rsbl.2015.0592
Roll, U., Feldman, A., Novosolov, M., Allison, A., Bauer, A. M., Bernard, R., Böhm, M., Castro-Herrera, F., Chirio, L., Collen, B., Colli, G. R., Dabool, L., Das, I., Doan, T. M., Grismer, L. L., Hoogmoed, M., Itescu, Y., Kraus, F., LeBreton, M., . . . Meiri, S. (2017). The global distribution of tetrapods reveals a need for targeted reptile conservation. Nature Ecology & Evolution, 1(11), 1677–1682. https://doi.org/10.1038/s41559-017-0332-2
Row, J. R., & Blouin-Demers, G. (2006). Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in milk snakes. Oecologia, 148(1), 1–11. https://doi.org/10.1007/s00442-005-0350-7
Seebacher, F & Franklin, C. E . (2003). Prostaglandins are important in thermoregulation of a reptile (Pogona vitticeps). Proceedings. Biological sciences / The Royal Society. 270 Suppl 1. S50-3. 10.1098/rsbl.2003.0007.
Smith, K. R., Cadena, V., Endler, J. A., Porter, W. P., Kearney, M. R., & Stuart-Fox, D. (2016). Colour change on different body regions provides thermal and signalling advantages in bearded dragon lizards. Proceedings of the Royal Society B: Biological Sciences, 283(1832), 20160626. https://doi.org/10.1098/rspb.2016.0626
Media Attributions
- Picture1
- Thermoregulation
- Thermoregulation-5
- Thermoregulation-8
- Thermoregulation-9
- Thermoregulation-10
- 4502_Thermoregulation.001
- Thermoregulation-7
- 4502_Thermoregulation.001
- Screenshot 2025-09-10 at 4.59.23 PM