10 Feeding
Dentition
Modern amphibians have pedicelate and biscupid teeth. The pedicel allows the teeth to bend inward (toward the inside of the mouth) but not outward which likely helps with the transport of struggling prey.

Turtles lack teeth entirely (they are edentate) but have a keratinous beak. Like for birds, the share of turtle’s beak varies with their diet. For instance, northern map turtles feed mainly on molluscs and have enlarged bony plates (called alveolar surfaces) on their upper and lower jaw that they use to crush molluscs. Snapping turtles are predators that often feed on prey much larger than themselves. They tear apart their prey a bit like a hawk would. the beak of snapping turtles is reminiscent of a bird of prey beak. It has sharp edges and a pointy hook. The jaws of some species of turtles bear serrations which function like teeth to slice through tough plant material.


In other reptiles, teeth are attached to the jaw bones in different ways. The type of teeth attachment to the jaw bone is called teeth implantation. The teeth of crocodilians sit in a socket and are attached to the jawbone with ligaments. This type of dentition is called thecodont. Mammals also have a thecodont dentition. In tuataras, teeth are attached to the crest of the jawbone and are rarely replaced. This type of dentition is called acrodont and is also found in some squamates including chameleons, agamid lizards, and some snakes. In most squamates, teeth are attached to the inner surface of the jawbone and are continually replaced. This type of dentition is called pleurodont. In some squamates including snakes, the base of the tooth is surrounded by some bony tissue; a dentition called subthecodont.

Many mammals including primates have a heterodont dentition meaning that a given species has different types of teeth (molars, pre-molars, incisors, canines) specialized for different functions such as piercing, slicing, and chewing. Reptiles do not show such level of teeth specialization within species and many species have a homodont dentition, meaning that all the teeth in the mouth have a similar morphology. Teeth specialization in reptiles is however present between species. Most reptiles are predators and thus have pointy teeth designed for piercing integument of their prey. Monitor lizards (family varanidae), which feed on large prey, have teeth with a sharper and serrated inner edge. Their teeth act as little steak knifes slicing through skin and muscles.

Most snake species swallow their prey whole and many feed on prey that are large relative to their body size (macrophagy). The dentition and skull of snakes reflect this feeding habit.

Snakes have several, backward facing teeth preventing a prey to wiggle out of their mouth. On the lower jaw, only the dentary bones bare teeth as in other squamates but on the upper jaw, teeth are found on the maxilla (as in other squamates) but also on the pterygoid and palatine bones.
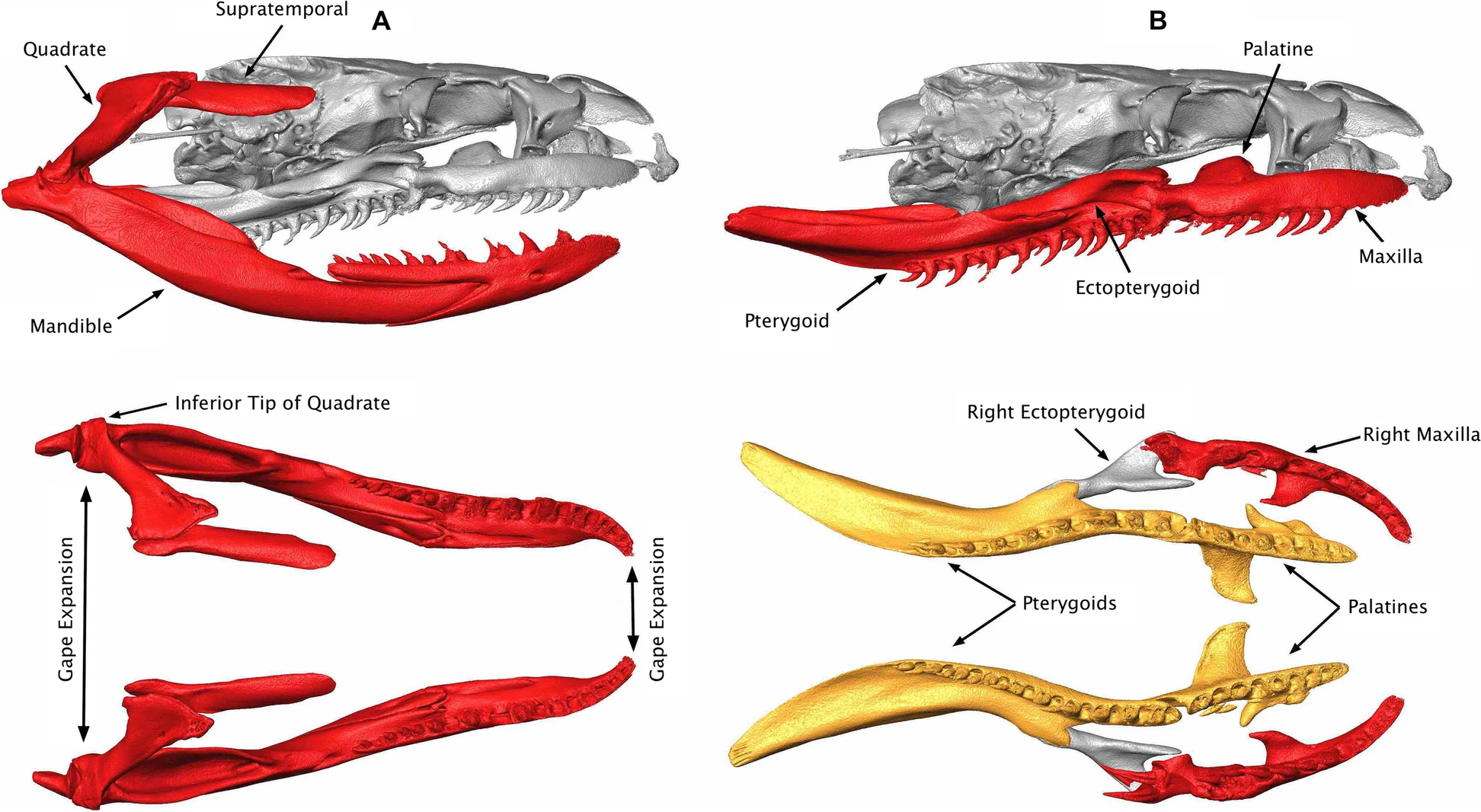
In some snakes, the teeth near the front of the mouth are somewhat S-shaped with the tip not as backward facing as in other teeth. This recurved shape facilitates the penetration of the teeth in the flesh of the prey when the mouth of the snake first contacts the prey during a strike. Once the tip of the tooth is embedded in tissues, it sinks deeper into it as the prey tries to escape.
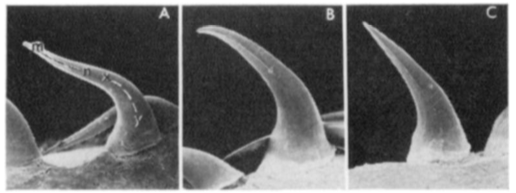
Although the vast majority of snakes species are not venomous, many species subdue their prey by envenomating them. All species in the families elapidea, viperidae, and atractaspididae are venomous. Some members of the family colubridae are also considered venomous. Venom is produced in the venom glands located in the cheek area. The venom glands are connected to fangs via ducts. The fangs are either grooved or hollow and inject venom in prey like hypodermic needles.
In viperids, the fangs are the only maxillary teeth present, and they are so long that they must fold backward when the snake closes its mouth. During a strike, the fangs unfold and rest perpendicularly to the upper jaw. Viperids and elapids are often called front-fanged snakes because their fangs (maxillary teeth) are at the front of the mouth. Other species of venomous snakes, such as the infamous boomslang (Dispholidus typus) are rear fanged. Their fangs are located further inside the mouth and are not hollow but grooved.


Skull Morphology
The muscles responsible for the closure of the jaw are called the jaw adductor muscles. In turtles, the enlarged middle ear cavity (a.k.a otic chamber; Ot. Ch. in figure below) which is formed mainly by the quadrate bone is in the way of these adductor muscles.The jaw adductor muscles of turtles are thus near the rear end of the skull. This position creates a mechanical problem for turtles because muscles generate the most force when they attach perpendicularly to a bone. Due to their backward position, the jaw adductor muscles of turtles, are parallel to the jaw bone. An analogy is helpful here. Imaging you are standing on a balcony and you need to pull up a bucket filled with water without spilling it. The bucket handle is attached to a rope. If the bucket is right under you, pulling it up without spilling is easy. In this analogy, you are the jaw muscles, the bucket is the jaw and the rope is the tendon attaching the muscles to the jaw. Now imaging the bucket is not just under the balcony but 30 meters away from it so the rope is no longer perpendicular to the bucket but somewhat parallel to it. If you pull on the rope, the bucket will fall on its side and the water will spill. This situation is similar to the problem faced by turtles. Their jaw adductor muscles are behind the lower jaw and the muscle fibers run parallel to it making jaw closure rather inefficient. The solution is a pulley. A pulley placed on top of the bucket makes the rope perpendicular to the bucket so it can be lifted without spilling. In turtles, the tendon attaching the muscles to the jaw pass through a kind of pulley and attach perpendicularly to the lower jaw. This pulley is formed by a projection of one of the skull bones. This adaptation evolved twice in each of the two sub-orders of turtles. In the cryptodyres, the pulley is formed by the quadrate bone. In the pleurodires, the pterygoid forms the pulley.

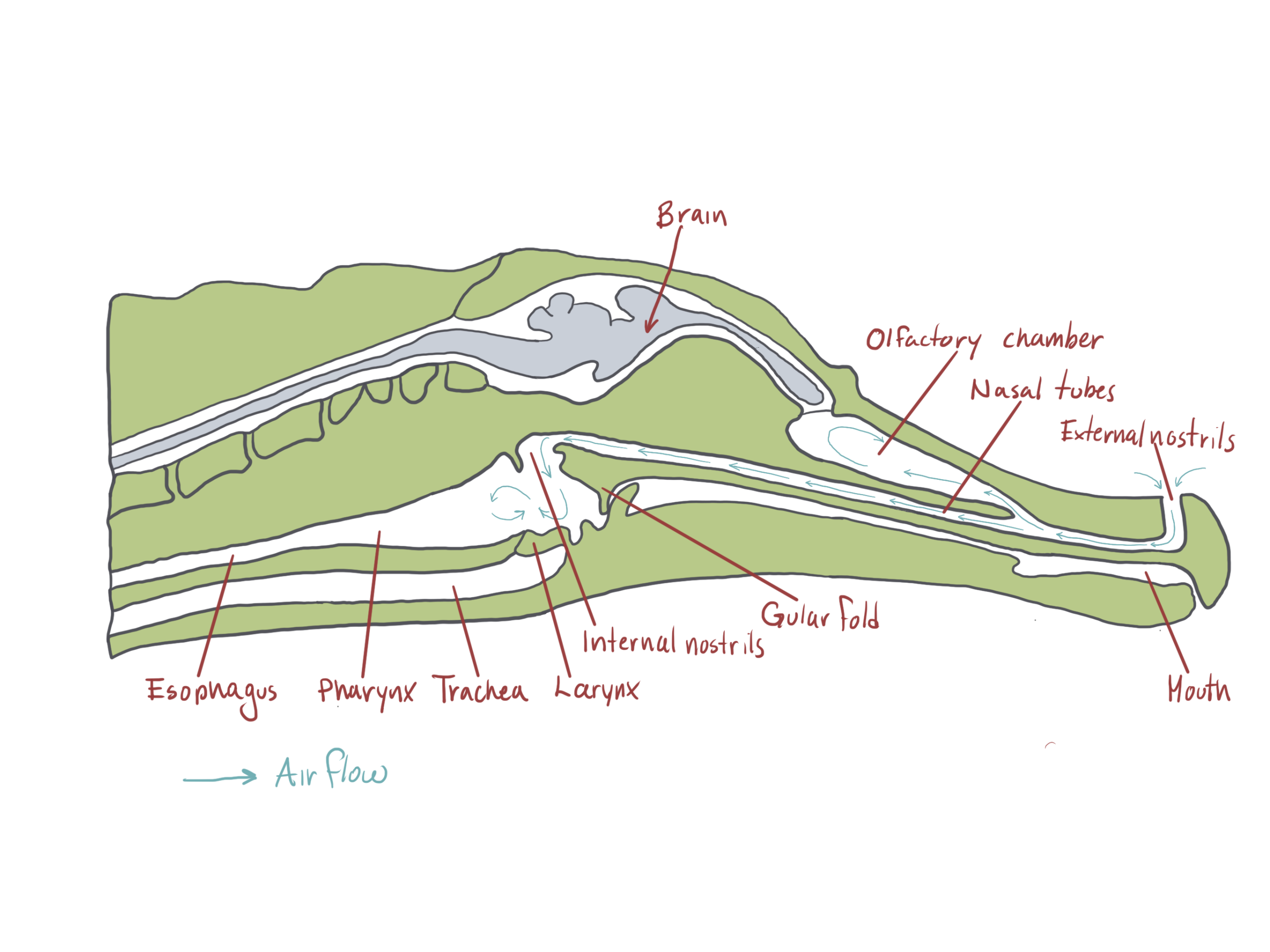
All modern crocodilians are aquatic predators which is reflected in their skull morphology. In other reptiles, the nasal passage opens in the roof the mouth but in crocodilians, the nasal passage is separated from the mouth by a bony palate, called a secondary palate. The secondary palate is formed by the enlargement of the maxilla, premaxilla and palatal bones. The internal nares (choane) open deep inside the mouth and can be shut by flap of skin. This flap keeps water from entering the nasal openings while crocodilians feed underwater. As we have seen in unit 3, crocodilians along with rhynchocephalians are the only living reptiles that are anatomically diapsid (skull with two temporal openings). The lower edge of the lower temporal opening is bounded by the lower temporal bar which adds stiffness to the skull. The persistence of the lower temporal bar may be adaptive as crocodilians exert a tremendous amount of force with their jaws when feeding.

Cranial Kinesis
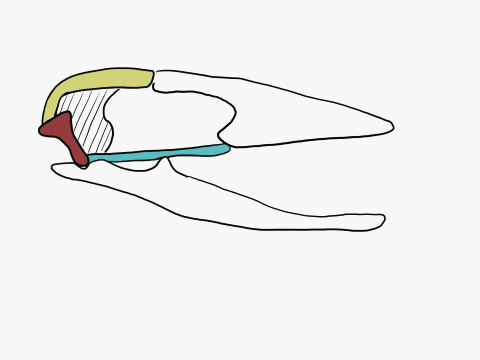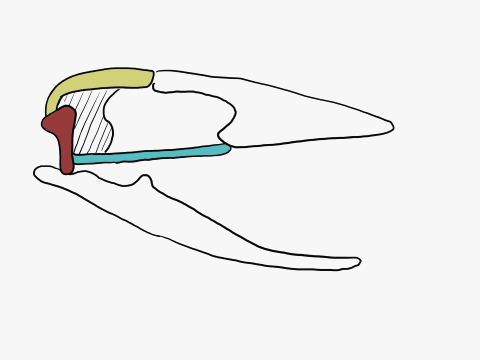
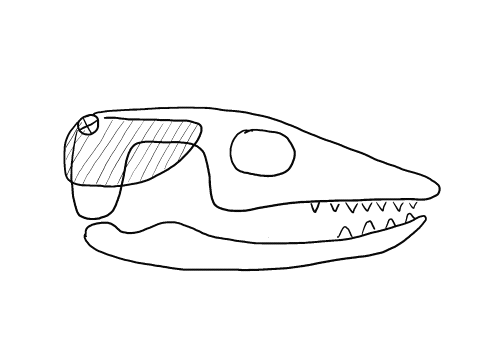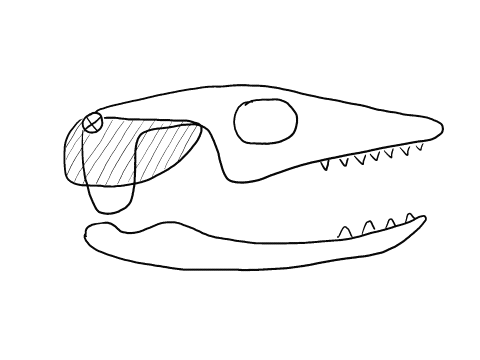
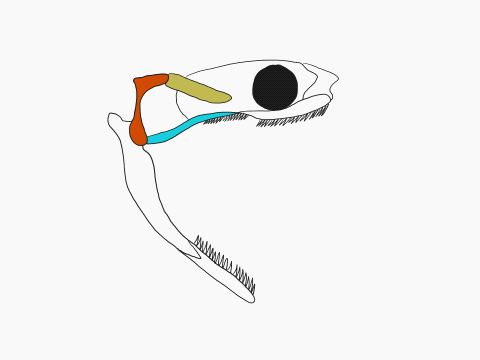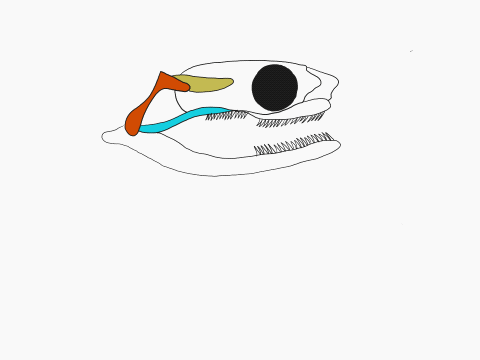
In contrast to crocodilians and rhyncocephalians, squamates have lost their lower temporal bar, freeing the quadrate (red) which can now articulate with the squamosal (yellow) and the pterygoid (teal) creating a form of cranial kinesis called streptostyly.
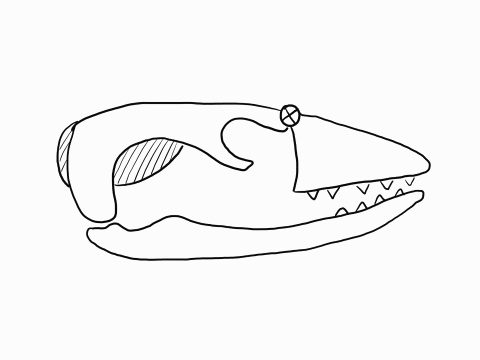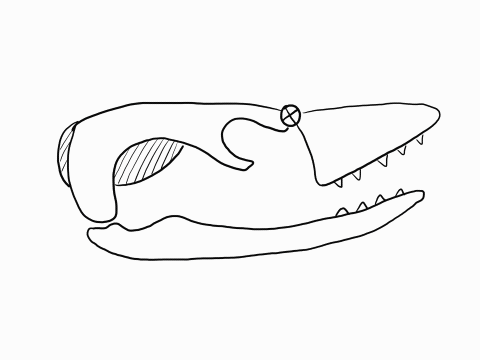
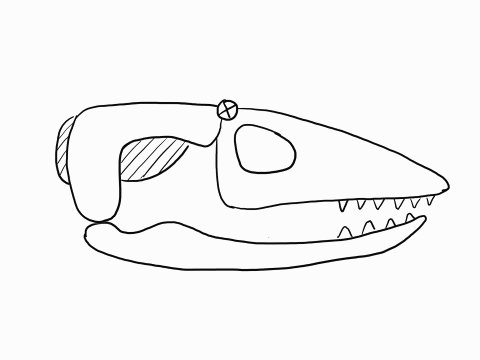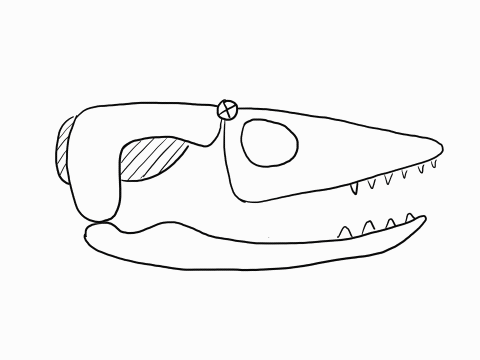
Other articulation points, called transcranial joints, may also be present in squamates: one at the rear end or the skull (metakinesis), one behind the orbit of the eyes (mesokinesis), and one in front of the orbit of the eyes (prokinesis). Most lizards have meta and mesokinetic joints in addition to streptostyly. Most snakes also display prokinesis. These joints allow the skull to change its shape during feeding.
 |
 |
 |
The kinetic skull of lizards can be viewed of as a frame with articulated corners. Pushing or pulling on one corner changes the shape of the frame. When the frame changes shape, the angles of the snout and teeth also change. This deformation of the skull keeps a downward pressure on the prey. If the skull was stiff, the jaws would close down on prey like scissors. If you try to cut a hard object like a small branch with scissors, it may slide out due to the force from the side as well as from the top and bottom. The deformation of the skull reduces the risk of a hard prey slipping out as the lizard chomps down on it.


Cranial kinesis in snakes and macrophagy
Snakes are famous for engulfing huge prey relative to their size but not all snakes use this feeding tactic. Most species of the basal lineages Scolecophidia feed on a large number of small prey. These snakes, like the blind snakes, Typhlopids and Leptotyphlopids, which specialize on ants and termites, have robust skulls with few teeth and a small gape. Some decapitate their prey to make them easier to ingest! One species of snake, Gerard’s water snake feeds mainly on crabs and evolved the ability to tear crabs apart to ingest smaller morsels. Most snakes however feed by ingesting few large prey whole; a feeding mode called macrophagy.

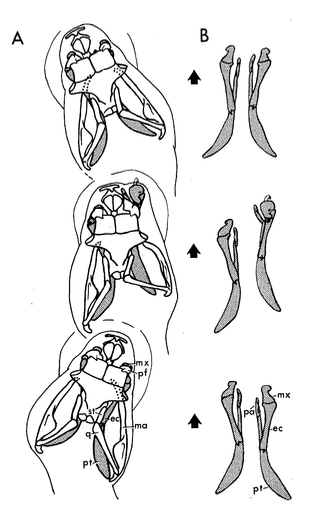
Cranial kinesis is essential for macrophagy and snakes have taken it to an extreme with as many as eight movable joints in their skulls. Snakes evolved from lizards. We’ve seen that lizards lost the lower temporal bar and thus retain only one temporal opening which permit streptostyly. Snakes lost the upper temporal bar as well further enabling streptostyly. The quadrate of snakes have more freedom of movement than in lizards. The quadrate articulates with the supratemporal which itself articulates with the brain case. These articulations enable the enlargement of the gape. Several other bones that, in other squamates are fused or immobile, can move independently from the brainscase in snakes.
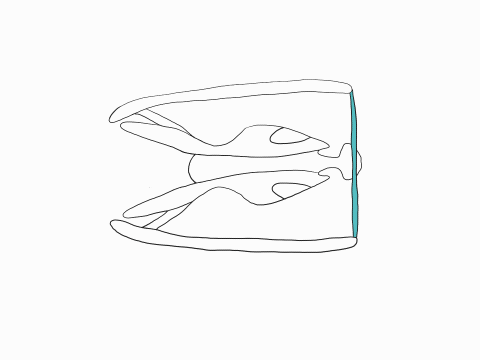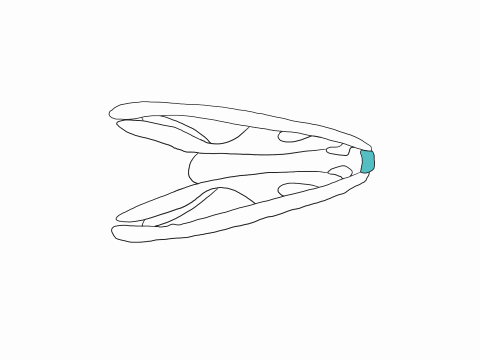
In all other vertebrates, the upper and lower jaws, are fuse at their anterior tips. In snakes the two halves of the jaws are attached by elastic ligaments allowing the two halves to spread apart and move independently from one another. Snakes can thus increase the gape of their mouth both vertically and horizontally.
A key adaption for macrophagy is the ability to independently move each side of the skull. This is possible due to the lost of bony joints between the upper jaw and the brain case. Instead, these two parts of the skulls are connected only by ligaments. These loose connections between the skull and the jaws allow snakes to perform the so-called pterygoid walk. In the pterygoid walk, snakes walk their jaws over the prey. During this process, one side of the upper jaw is lifted and moved forward while the other side and the lower jaw hold the prey in place. By alternating sides, the snake moves its head forward and around the prey. The brain case of snakes lacks the meta and mesokinetic joints seen in lizards probably because the braincase is the main anchor points for the jaw muscles, and must thus be stiff. The solid brain case also protects the brain from potential harm caused by fighting prey. Snakes however have a prokinetic joint and some species can also move the tip of their snout independently through a rhinokinetic joint.

Suspension feeding
Suspension feeding (a.k.a filter feeding) involves filtering out small food morsels suspended in the water column (plankton). In some cases, the animal may first scrap a surface to put the food particles in suspension before filtering them out of the water. Among amphibians and reptiles, tadpoles are the only suspension feeders. Tadpoles are primarily herbivorous and feed on periphyton and phytoplankton. The mouth of a tadpole consists of a keratinized beak siting at the center of a flat disc called the oral disc. The margin of the oral disc bears sensory papillae. The oral disc is also ornamented with rows of keratinized projections called labial teeth which are used to scrap the periphyton from the substrate. The configuration of the labial teeth is one of the main criteria used to identify tadpoles to the species.

Tadpoles use their mouth and labial teeth to scrap and put the periphyton attached to various surfaces suspension. The suspended particles are then sucked inside the mouth using the pumping action of their respiratory system. The food particles are filtered out of the water before the water reaches the gills. A membrane, called the ventral velum, divides the mouth cavity from the pharyngeal cavity where the gills reside. The ventral velum is lined with a sticky mucus to which the larger food particles stick. The smaller particles that escape the velum’s sticky trap are intercepted by a fine branchial filter associated with the gills. All captured food particles are coated with mucus and sent to the oesophagus, and the digestive tract.

As in many filter feeders, the feeding system of tadpoles is closely associated with gill ventilation. The water ventilating the gills also goes over the ventral velum and through the brachial filter. Tadpoles suck water in their mouth and nostrils (nares) by lowering the floor of their mouth cavity to increases its volume decreases its internal pressure which causes water to flow in through the mouth and nares. The floor of the mouth is then raised to push the water over the ventral velum and into the pharyngeal cavity where it flows through the brachial filter and over the gills. While the floor of the mouth is raised, the mouth and nares are shut to prevent the water from escaping through those openings. After having gone over the ventral velum, the brachial filter, and the gills, the filtered and de-oxygenated water exits through the spiracle.


Suction Feeding
Suction feeding involves sucking in water along with whatever prey in contains inside the mouth. This feeding mode is common in aquatic turtles and salamanders. One obvious challenge with suction feeding is dealing with the large volume of water entering the mouth. Larval and neotenic salamanders have gills and gill openings so the water entering the mouth can exit through these openings (as in fish).

Adult salamanders and turtles lack gill openings so the water entering the mouth must exit by the mouth. These suction feeders thus tend to have a large and stretchy throat to accomodate a large volume of water. Humpback whales ramming schools of fish with their mouth wide open have the same problem and they too have a very stretchy throat. In the mata mata (turtle) and the Japanese giant salamander slow motion videos below, note how much the throat area expands during feeding.


The key behind suction feeding is to create a vacuum inside the mouth at lightning speed. This is achieved by sucking water in a large cavity through a small opening. Think of a syringe. The smaller the opening relative to the body of the syringe, the faster the water will flow in when the plunger is pulled. Suction feeding function in a similar way. A large cavity is created inside the mouth but the opening of the mouth itself remains relatively small.
Suction feeders are capable of rapidly expanding their buccal cavity which is how the vacuum sucking the water in is created. Recall from our lecture on gas exchange that the hyoid apparatus is used to increase the volume of the buccal cavity and plays an important role in the positive pressure ventilation system of amphibians. The ability of the hyoid to increase the volume of the buccal cavity is also useful during suction feeding and suction feeders usually have a large hyoid apparatus.
An extreme example of suction feeding in reptiles is the bizarre Mata mata turtle (Chelus fimbriatus) from South America which can suck-in a fish 2 cm away in 0.04ms. The Mata mata has a massive hyoid apparatus and a highly expandable pharynx and esophagus that can engulf a large volume of water. Look at the 3D rendition of a Mata mata skull below to see how large the hyoid apparatus is. The bones labelled 3 to 8 are part of the hyoid apparatus. During feeding, the hyoid is rapidly depressed to increase the volume of the mouth cavity. The tongue does not play an important role in suction feeding and it is greatly reduced in the Mata mata. A small tongue increases the volume available for water to flow in and reduces resistance as water flows in. Other species of turtles also employ suction feeding to some extent including the common snapping turtles and the common musk turtle.

Suction feeding is well developed in aquatic salamanders. In salamanders with gills (larval and neotenic) water can exit through the gill openings (as in fish). The hyoid apparatus also plays an important role in increasing the volume of the oral cavity. Salamanders also raise their skull during suction feeding which further increase the volume of the mouth cavity. Meanwhile, the opening of the mouth remains relatively small, creating a strong suction drawing water in as well as the prey suspended in it. The water then exits through the pharyngeal slits which are too small for the prey to exit by.
Projectile feeding
Some amphibians and reptiles capture prey by shooting their sticky tongue at them. This mode of feeding is called projectile feeding. Projectile feeding takes advantage of elastic recoil to generate the force necessary to shoot the tongue outside the mouth. A familiar example of an elastic recoil system is a bow and arrow. When shooting an arrow with a bow, your arm muscles are not throwing the arrow but they stretch the elastic string to store potential energy in it. When you let go of the string, the potential energy is converted to kinetic energy that propels the arrow. The recoil of the string propels the arrow faster and further than you possibly can by throwing it using your arm muscles. Projectile feeders use an analogous mechanism to shoot their tongue outside of their mouth. They contract muscles to store potential energy in elastic tissues. When the stretched out elastic tissues let go, the tongue shoots out. This kind of system has two main advantages. First, it generates more power than muscle contractions alone can. Second, it is relatively unaffected by temperature. Indeed, while the speed of muscle contractions is affected by temperature, the speed of elastic recoil is not. This allows projectile feeders to feed successfully even at relatively low temperatures.

Many anurans including true frogs (Ranidae) and toads (Bufonidae) use a form of projectile feeding that resemble the action of a catapult. The tongue of frogs and toads is attached near the front of the mouth. To catch a prey, the tongue “flips out” of the mouth and slaps the prey with great force and speed (up to 270cm/sec). As the tongue flips out, it also stretches substantially (up to 200% its resting length) because of inertia (inertial elongation). The rapid movement of the tongue and its inertial elongation occurs because the lower jaw is rapidly depressed. The depression of the jaw appears to be powered in part by the recoil of elastic tissues. Indeed, milliseconds before the lower jaw is depressed and the mouth opened, the large jaw depressor muscles contract to stretch elastic tissues connecting the muscles to the jaw. This contraction stores potential energy in the stretched elastic tissues (like arming a catapult or turning the crank in a jack in the box). When the mouth starts to open, the tensed elastic tissues let go and the jaw depresses rapidly. Other muscles are also involved in the “flipping” of the tongue. The genioglossus medialis attaches the tongue to the tip of the lower jaw (see image below). When it contracts, it pulls the tongue forward. Another muscle, the submentalis, bulges as the genioglossus medialis pulls the tongue. The contraction of the submentalis helps the tongue pivot over the tip of the jaw. See animation below


The most extreme examples of projectile feeding are found in some plethodontid salamanders and in chameleons. As in frogs, the projectile mechanism depends to some extent on elastic recoil. Both group have incorporated part of their hyoid inside their tongue making what is essentially a tongue skeleton. In projectile feeding plethodontids (e.g. Bolitoglossa sp, Hydromantes sp), the hyoid has become integrated into the tongue so the salamanders not only shoot their tongue outside of their mouth, but also part of their hyoid apparatus! These salamanders can project their tongue up to 80% of their body length.


The hyoid apparatus of these salamanders includes a pair of long branches (epibrachial) extending into the abdominal cavity. These branches are wrapped by a tongue projecting muscles. When these muscles contract, the branches slide forward and come together. The anterior part of the hyoid comes out of the mouth with the tongue. Tongue retraction is achieved through the contraction of a pair of tongue retraction muscles which have their origin on the pelvis!
Chameleons can shoot their tongue nearly 200% of their body length. Two muscles make up the bulk of a chameleon’s tongue: a long retractor muscle that brings the tongue back into the mouth, and a powerful accelerator muscle located in front the retractor muscle that shoots the tongue outside the mouth. Both muscles wrap around a long rod derived from the hyoid called the entoglossal process. Between the muscles and the entoglossal process, lies several sheaths of elastic collagen. The fibers of the accelerator muscles run perpendicularly to the entoglossal process so when the muscle contracts, it stretches the underlying sheaths of collagen which store potential energy (iii in diagram). All of this happens millisecond before the tongue is launched (the equivalent of stretching the string on a bow). The tongue shoots out when the accelerator muscle and collagen sheath are pushed over the tip of the entoglossal process by the retractor muscle. At that point the elastic energy stored in the collagen sheaths is released and the accelerator muscle shoots out of the mouth towards the prey. The elastic energy stored in the collagen sheaths generates ten times more power than the accelerator muscle can.


Media Attributions
- Untitled.001
- Predation & Scavenging-32
- 12862_2023_2157_Fig3_HTML
- Screenshot 2025-10-28 at 2.04.39 PM
- Crocs_Palate
- Streptostyly 2
- Prokinesis 2
- Mesokinesis 2
- Metakinesis 2
- JEB198291.MovieS1
- Untitled_Artwork 6
- Untitled_Artwork 7
- Screenshot 2025-10-28 at 3.23.51 PM
- jez70028-sup-0002-sm1_gold1_free_prey_seq1_filtered_labeled(1)