6 Surviving the winter
Winter’s coming
Aside from sea turtles, reptiles and amphibians cannot undertake long migrations to escape winter. When winter arrives, amphibians and reptiles in Canada stay put and deal with the cold. The biggest challenge they face is keeping their cells from freezing. Unlike birds and mammals protect their cells from freezing keep by maintaining a high body temperature. Herps cannot generate enough metabolic heat to protect their cells from the assaults of ice. Ice crystals can puncture cell membranes, but the main issue with freezing is desiccation. Cells bathe in extracellular fluids and must regulate the flow of solutes through their semi-permeable membrane. As the extracellular fluid freezes, the concentration of solute increases causing water to leave cells. To protect their cells from the perils of freezing, animals have evolved a wide range of physiological, biochemical, and behavioural adaptations.

Amphibians and reptiles can’t remain (fully) active in winter (at high latitudes) because temperatures are too low for them to move about and perform other physiological functions (see thermoregulation). There is one notable exception however: The Mudpuppy (Necturus maculosus). This large neotenic, and thus fully aquatic, salamander lives in streams, rivers, and lakes and it remains active all winter. In fact, mudpuppies appear to be primarily active in winter. Mudpuppies feed on small fish, aquatic invertebrates and are sometimes captured by ice fishing adepts. Water snakes and wading birds like herons are the main predators of mudpuppies. It is possible that this species evolved to be active in winter to avoid these predators. Very little is known about the physiological adaptations allowing mudpuppies to remain active at temperature at which most herps become lethargic.
Amphibians and reptiles use three strategies to avoid lethal frost bites in winter. They can escape freezing (freeze avoidance) temperatures altogether by moving underground below the frost line. Another place to escape freezing is in water. Some streams and rivers do not freeze due to their high flow. The surface of lakes, and wetlands typically freezes in winter but the water below the ice remains liquid. Another strategy is to use physiological tricks to prevent bodily fluid to freeze at sub-zero temperatures. This strategy is called freeze resistance. A third strategy is to allow the extracellular fluid to freeze in a controlled manner but to prevent the intra-cellular fluid to freeze. This strategy is called freeze tolerance.
Freeze avoidance
Frost only penetrates the ground only to a certain depth called the frost line. Around Ottawa the frost line is about 1m deep. Animals can escape frost by going below the frost line. In Ontario, snakes, and terrestrial salamanders use this strategy to survive the winter. Toads like the American toad and the Fowler’s toad also use this strategy.

Snakes commonly overwinter with other snakes in dens called communal hibernacula. The largest aggregation of snakes in the world is in Manitoba where thousands of red-sided garter snakes (Thamnophis sirtalis parietalis) spend the winter together in communal dens. The tendency for snakes to overwinter in groups may reflect the limited availability of suitable hibernacula in northern landscapes. Most species of snakes cannot dig their own burrow but rather use existing burrows or cavities. Radio-telemetry work on Gray Rat Snake in Ontario showed that individual snakes will use the same hibernaculum year after year and some will travel long distances to reach them. Identifying and protecting communal hibernacula may thus be necessary for the conservation of snakes in some areas.

Lakes, ponds, streams, and rivers also offer frost free refugia in the winter and turtles as well as many species of frogs use these habitats to spend the winter. While some lotic water bodies (e.g. streams and rivers) may remain frost free all winter, the surface of most lentic water bodies (e.g. lakes and ponds) does freeze.

The fact that lakes and ponds of a certain depth do not freeze all the way to the bottom is somewhat counter-intuitive. You probably know that deeper you go in a lake, the colder the water gets, so why don’t lake freeze from the bottom up rather than the top down? The reason is a chemical quirk of water. As water cools down, its density increases but the relationship between density and temperature breaks down at 4°C. Water at 4°C is denser than water at any other temperature. That’s the quirk. Without that quirk, lakes would freeze from the bottom up, not the top down and life in them in winter would be impossible.
In summer, the water at the surface of the water is at its warmest but it cools down in the fall as the air temperature drops. The surface water eventually becomes colder than the water below it. When the surface water reaches 4°C, it sinks because it is heavier that the warmer water below it. As the denser surface water sinks, it pushes up the water below it to the surface. This phenomenon is called the fall lake turnover. As the turnover wheel spins, more water is brought up to the surface, cooled down to 4°C and then sink. Eventually, the water temperature will at 4°C from the surface to the bottom. At that point, the surface water keeps cooling down as the air temperature drops until it freezes. The ice becomes thicker as the winter progress but the lake never freezes down to the bottom. So in winter the water near the bottom is at around 4°C while the water near the surface is just above 0°C as shown in the graph below.

However, overwintering underwater poses a problem for gas exchange. Lungs obviously don’t work underwater so frogs and turtles brumating in water must rely on extra-pulmonary routes to get oxygen and excrete carbon dioxide. The primary strategy in both amphibians and reptiles is cutaneous respiration (see section metabolism and gas exchange).
Oxygen enters the water through either surface agitation or via the photosynthesis of aquatic plants and algae. Streams and rivers are typically well oxygenated because of the constant surface agitation. In winter, surface agitation prevents, or at least limits, the formation of ice. Streams and rivers thus have a steady supply of atmospheric oxygen in winter. For species overwintering in streams and rivers, such as the wood turtle (Glyptemys insculpta) oxygen availability may thus not be a major issue.
Lakes and ponds are a different story, however. The ice forming at the surface of ponds and lakes cuts off the water body from the atmosphere so oxygen ceases to enter by diffusion. The only input of oxygen in a frozen lake is photosynthesis which is minimal in winter due to cold water temperature and shading by ice and snow. Moreover, aquatic organisms including bacteria continue to consume oxygen in winter so the concentration of oxygen in water declines as winter progresses. Overwintering in such conditions can be a challenge.

If the concentration of dissolve oxygen drops to low levels (hypoxia) or if the water becomes depleted in oxygen (anoxia), frogs and turtles must switch from aerobic to anaerobic metabolism. Most species of frogs and turtles however cannot survive an entire winter by relying on anaerobic metabolism. For instance, the spiny softshell turtle can only survive 14 days submerged in anoxic water at 3°C. The common musk turtle can last 22 days and the northern map turtle about 50 days. Although impressive for tetrapods, these feats are far from the duration of the ice-cover in our part of the world. For instance, Lake Opinicon, just south of Ottawa, is covered with ice for 95 to 155 days every year. The painted turtle and the common snapping turtles are notable exceptions with regards to their tolerance to anoxia. Both species can spend entire winters without oxygen. Snapping turtles can spend 118 days submerged in anoxic water at 3°C but painted turtles are the real champs and can last more 150 days under the same conditions!
The overwintering physiology of painted turtles has been studied in great details. The book Life in a Shell by Donald Jackson offers an excellent summary of this research and other amazing physiological feat turtles are capable of. To survive for months without oxygen painted turtles reduce their metabolism to an extremely low level and buffer the lactic acid produced by glycolysis.
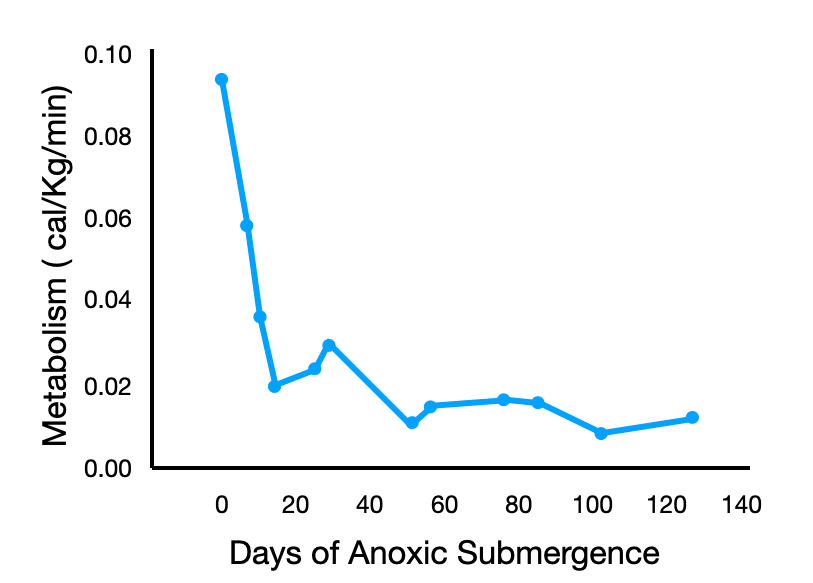
The metabolism of ectotherms varies as a function of their body temperature. In reptiles, a drop of 10°C in body temperature typically leads to a two to three folds decrease in metabolism. The body temperature of a brumating turtle hovers just above the freezing point so metabolic rate is greatly reduced by the effect of temperature alone but painted turtles can reduce their metabolism beyond the effect of temperature alone. When a painted turtle is moved from normoxic to anoxic water at the same temperature (3°C), its metabolism drops by 90% in a few days. This metabolic drop is called metabolic depression. Metabolic depression appears to be achieved in part by slowing down two ATP hungry processes: ions pumping by cells, and protein synthesis.
Ions channels are proteins embedded in cell membranes through which ions move passively. These channels can open and close. When the channels are open, sodium ions leak into the cell so the cell has pump these ions back out using the sodium potassium pump. This active transport maintains the chemical and electrical gradients across the cell membrane. Pumping ions against their concentration gradient consumes a lot of ATP however. Anoxic turtles slow down ion pumping by reducing the number of ion channels in their cells and by keeping the remaining ones closed more often; a phenomenon called channel arrest. Less ions leak in, so less ions must be pumped out, so less ATP is consumed. Of course, channel arrest impedes the ability of cells to do their job, but hibernating painted turtles are minimally active so they can get away with it.
Thanks to metabolic depression, a cold and anoxic painted turtle consumes about 10 000 times less energy than a similar size mammal! Painted turtles can last for 150 days on about 1000 calories; the equivalent of two 100g Snicker chocolate bars.
The second strategy consists of buffering the lactic acid produced by the anaerobic metabolism. Overwintering painted turtles do not need much ATP but they still need some. In the absence of oxygen, ATP comes from glycolysis. As you know, lactate (CH3CH(OH)COOH) is a by-product of glycolysis. In solution, lactate dissociates into lactate ions (CH3CH(OH)COO–) and H+ ions creating lactic acid. The accumulation of lactic acid can disrupt normal physiological functions and cause death. This occurs in turtles when the plasma pH falls below 7.2. Plasma acidification is what kills most species of frogs and turtles exposed to anoxic water in winter. A normal concentration of lactate for a human and a turtle (under normoxic conditions) is about 1mmol/L. In an anoxic painted turtle, the concentration of lactate in the plasma reaches 200 mmol/L! For reference, if you put an athlete on a treadmill and make them run until they collapse from exhaustion, the concentration of lactate in their plasma would be about 25 mmol/L. Amazingly, anoxic painted turtles manage to maintain a nearly neutral pH despite producing enormous amounts of lactate (see graph below). The secret of their success is their skeleton.
Painted turtles take advantage of their voluminous skeleton to buffer their pH during anoxic hibernation. Firstly, they sequester much of the lactate in their shell in a still unknown neutralized form. Secondly, they use the calcium carbonate from their bones as a buffer. The calcium carbornate dissociates into calcium and carbonate ions. The carbonate ions scavenge the hydrogen ions released by the lactate and neutralize the blood pH. In essence painted turtles use their skeleton the way we used calcium based anti-acid tablets to calm heart burn.

Using these two strategies, Painted Turtles maintain a nearly neutral pH for months while relying entirely on glycolysis. Under similar anoxic conditions, other species (except for common snapping turtles) experience acidosis and die after 14 to 50 days. These so-called anoxia intolerant species rely on a steady supply of oxygen in the winter. Interestingly, some species like the northern map turtle and the spiny softshell turtle brumate communally. The proximate causes of communal hibernation in turtles are not clear but may indicate that suitable sites with enough dissolved oxygen are limited.

Freeze Resistance
In temperate areas turtles lay their eggs in the early summer and the eggs hatch in the late summer and early fall. However, in some species including Painted Turtles and Northern Map Turtles, most hatchlings do not leave the nest when they hatch. They instead remain in the nest through the fall and winter and emerge in the spring. This phenomenon is called delayed emergence and its evolutionary causes are not well understood.

Species delaying their emergence from the nest until the spring experience freezing temperatures because nests are above the frost line. For instance, nests of northern map turtles in Kingston, Ontario will go down to -8°C. Yet the hatchlings inside the nests manage to keep their bodily fluids liquid at such a low temperature. Two strategies make this possible. Firstly, hatchling turtles lower the freezing point of their extracellular fluid by increasing its solute concentration by accumulating solutes such as sugars and urea. This strategy however is insufficient to drop the freezing low enough for hatchlings to survive all winter at temoeratures as low as -6 or 7°C. The second strategy hatchlings use is supercooling.
For a liquid to freeze, ice crystals must organize themselves around a surface. These ice forming surfaces are called ice nucleating agents and may be impurities or existing ice crystals. Turtle overwintering in the nest void their guts of debris that could act as ice nucleating agents. Some species such as the Painted Turtle also sport an extra lipid layer on its skin which acts as a barrier preventing ice crystals from penetrating their tissue and hence trigger freezing. Interestingly, hatchlings of common snapping turtles do not overwinter in their nest, and they lack this lipid layer. Supercooling is a dangerous business however. A supercooled liquid is unstable and can freeze in a split second if a nucleation site becomes available or if temperature drops too low. The risk of freezing increases with the duration of exposure to freezing temperature as well as with the minimum temperature experienced.

Freezing Tolerance
The third and most extreme strategy used by herps to survive the winter is allow their extracellular fluid to freeze while keeping the intracellular fluid liquid. This strategy is called freeze tolerance and it is most developed in frogs but was also documented in some salamanders, snakes, and lizards, as well as in hatchling painted turtles. In Ontario, the wood frog (Lithobates sylvatica), the spring peeper (Pseudacris crucifer), the chorus frog (Pseudacris triseriata/maculata), the gray tree frog (Hyla versicolor), as well as hatchling painted turtles use freeze tolerance to survive the winter. Most of the work on the physiology and biochemistry of freeze tolerance has been conducted on wood frogs in the laboratory of Dr. Kenneth Storey here at Carleton.
Freeze tolerance involves a complex cascade of biochemical and physiological changes that are beyond the scope of this course (see this paper by Storey & Storey for a recent review) but we will summarize some of the key aspects here.
As we’ve seen above, supercooled liquids risk freezing when nucleation sites become available. Moreover, ice crystals can cause mechanical damage to cell membranes. Freeze tolerant species allow their extracellular fluid to freeze but in a highly control manner. Ice binding proteins act as nucleation sites at temperature just below the freezing point. These proteins not only initiate freezing in a controlled manner but also limit the size of ice crystals. As the extracellular fluid freezes, its solute concentration increases, and water is drawn out of the cell. To keep their integrity, the cells limit the outflow of water by increasing the internal solute concentration with osmolytes such as glucose, glycerol, and urea. These osmolytes reduce the osmotic gradient between the intra and extracellular fluid and limit how much water exits the cell. The osmolytes also reduce the freezing point of the intracellular fluid (cryoprotectants).
Freeze tolerant species rely on glycolysis when in their frozen state. As we’ve seen above, prolonged glycolysis can lead to lethal acidosis. Freeze tolerant species reduce their ATP consumption by depressing their metabolism. They do so by reducing gene transcription, and protein synthesis, by increasing the longevity of enzymes, and by the selective atrophy of certain tissues and organs.

References
Packard, Gary & Packard, M.J. (2001). The overwintering strategy of hatchling painted turtles, or how to survive in the cold without freezing. BioScience. 51. 199-207.
Jackson, Donald. (2002). Hibernating without Oxygen: Physiological Adaptations of the Painted Turtle. The Journal of physiology. 543. 731-7. 10.1113/jphysiol.2002.024729.
Storey, K & Storey, J. (2017). Molecular Physiology of Freeze Tolerance in Vertebrates. Physiological Reviews. 97. 623-665. 10.1152/physrev.00016.2016.
Ultsch, Gordon. (2006). The ecology of overwintering among turtles: Where turtles overwinter and its consequences. Biological reviews of the Cambridge Philosophical Society. 81. 339-67. 10.1017/S1464793106007032.
Weatherhead, PJ . (2011). Temporal and thermal aspects of hibernation of black rat snakes (Elaphe obsoleta) Ontario. Canadian Journal of Zoology. 67. 2332-2335. 10.1139/z89-328.
Media Attributions
- Untitled.001
- Untitled.001
- Screenshot 2025-10-01 at 11.09.23 AM