1.3 Mechanisms of Evolution
Many students mistakenly use the term “evolution” as if it means the same thing as “natural selection,” but they are not identical. Evolution refers to the overall process of change in the genetic makeup of populations over time. Natural selection is just one of the mechanisms that drive evolution. It occurs when individuals with traits better suited to their environment are more likely to survive and reproduce. It is the only way in which populations evolve adaptively.
We can see more random changes in a population through other evolutionary mechanisms, such as random mutations, genetic drift, and gene flow.
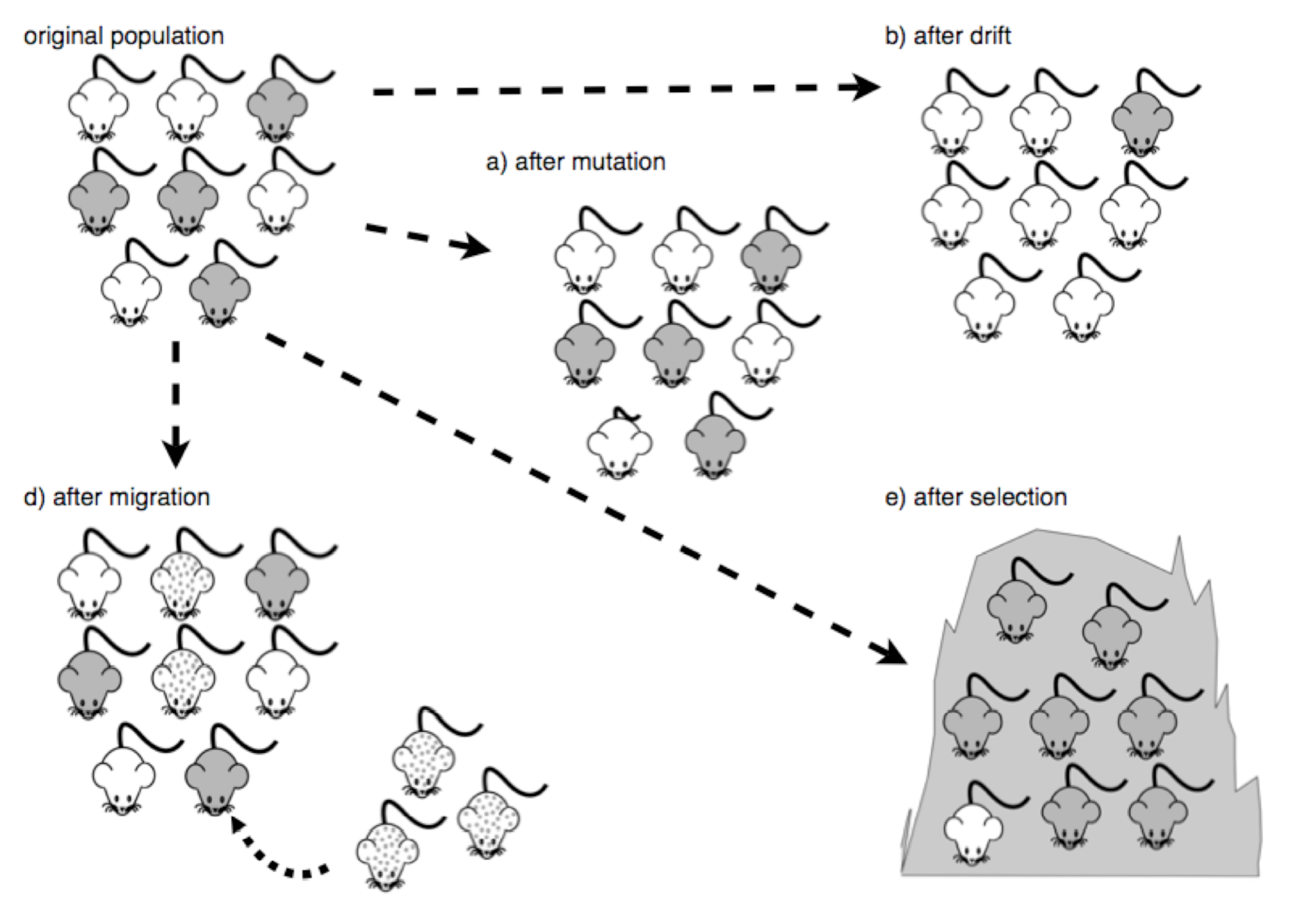
Mutations
A mutation is a random change in the DNA sequence of an organism. Errors in DNA replication and exposure to environmental factors (like radiation or chemicals) can lead to mutations.
A population has high genetic variation when there are many different variants of genes within the population. Mutations contribute to this variation by creating new gene variants. In this sense, mutations are the ultimate source of genetic diversity in evolving populations and provide the raw material for natural selection to act upon.
When a novel mutation arises, the population has changed, so mutation alone is a mechanism of evolution. However, a single random mutation is unlikely to have a major impact on its own. Other evolutionary mechanisms—such as natural selection, gene flow, and genetic drift— play a larger role in determining whether that mutation becomes widespread.
Mutations can be harmful, beneficial, or neutral, depending entirely on the environmental context. Harmful mutations may reduce an individual’s chances of survival or reproductive success and are typically removed from the population through natural selection. Beneficial mutations, on the other hand, can provide an advantage in a specific environment, such as improving survival or reproduction, and are more likely to spread. These mutations are often associated with natural selection because they lead to adaptations that help organisms respond to selective pressures. Some mutations have no effect on an organism’s fitness. These neutral mutations are not influenced by natural selection and instead change in frequency due to other evolutionary mechanisms like genetic drift or gene flow.
Genetic Drift
Genetic drift is a random change in allele frequencies in a population, especially noticeable in small populations. Unlike natural selection, which is driven by advantages in survival or reproduction, genetic drift happens by chance. For example, if an individual dies before reproducing or fails to find a mate, their alleles may be lost from the population, not because they were harmful, but just by luck.
In small populations, losing even one individual can significantly change the gene pool. For instance, in a group of 10 individuals, one death could remove 10% of the genetic variation. In a population of 1000, that one individual represents only 0.1% of the overall gene pool; therefore, it has much less impact on the population’s genetic structure and is unlikely to remove all copies of even a relatively rare allele.

There are two key types of genetic drift:
Bottleneck Effect
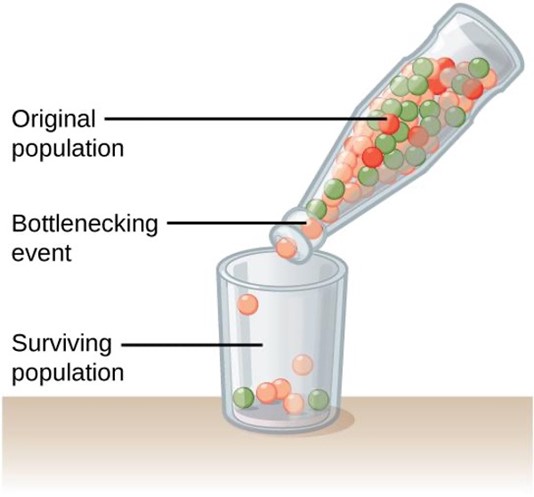
Natural events, such as an earthquake disaster that randomly kills a large portion of the population, can magnify the effects of genetic drift. These situations, where a large portion of the gene pool is suddenly and randomly eliminated, are called the bottleneck effect (Figure 1.3.2). At once, the survivors’ genetic structure (i.e., allele frequencies) becomes the entire population’s genetic structure, which may be very different from the pre-disaster population. Almost certainly, rare alleles are lost from the population, resulting in a decrease in the variability in the population. The survivors reproduce to create the next generation, which can have dramatically different allele frequencies compared to the population before the bottleneck event occurred. Bottleneck effects are a special case of genetic drift where the effects of drift are particularly strong because a population’s size has been dramatically reduced.
Founder Effect
- Another scenario in which populations might experience a strong influence of genetic drift is if some portion of the population leaves to start a new population in a new location. It is likely that the small number of individuals who found the new population have different allele frequencies than the entire population due to random chance. Thus, the new population can end up having very different allele frequencies than the original source population due to genetic drift – this is called the founder effect.
Tristan da Cunha, one of the most remote inhabited islands in the world, provides a classic example of the founder effect. In 1816, the island was settled by just a few individuals, including one man who unknowingly carried a rare allele for retinitis pigmentosa, a genetic disorder that causes progressive blindness. Due to the island’s isolation and small population size, this allele was passed down through generations and became unusually common. Among roughly 240 descendants, several individuals developed blindness, and others were carriers of the condition. This high frequency of a rare genetic disorder, far above global averages, occurred not because the allele was advantageous, but purely due to chance (Berry, 1967).
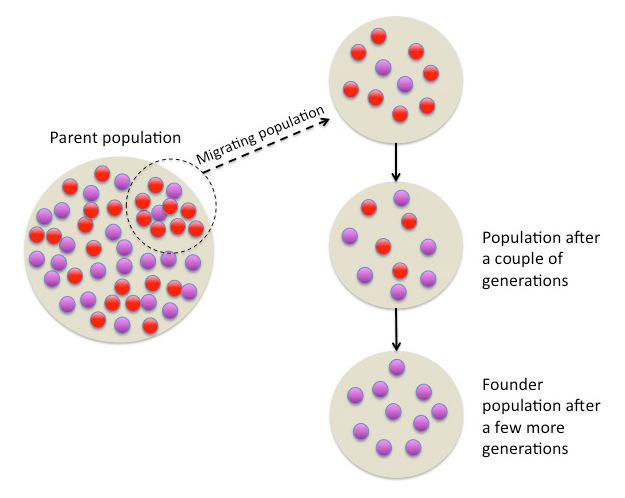
Figure 1.3.4 Image Description
The image is a diagram illustrating the founder effect in population genetics.
- On the left, a large beige circle labelled “Parent population” contains many individuals represented by small red and purple circles mixed together.
- A smaller group of circles, highlighted within a dashed oval, is labelled “Migrating population” and shown moving to the right with a dashed arrow. This migrating group contains a higher proportion of red circles than in the original parent population.
- To the right, the migrating group forms a new, smaller beige circle representing the initial population in the new location. This group has fewer total individuals, mostly red with some purple.
- An arrow points downward to the next beige circle labelled “Population after a couple of generations”, showing more mixing but still a smaller, less diverse gene pool compared to the original population.
- A final arrow leads to the bottom beige circle labelled “Founder population after a few more generations”, which now consists almost entirely of purple circles, showing reduced genetic variation over time due to genetic drift.
The diagram visually explains how a small migrating population can lead to reduced genetic diversity and altered allele frequencies in future generations.
Genetic drift can lead to significant evolutionary changes over time, especially in small populations where random events have a larger impact.
Gene Flow
Gene flow refers to the flow of alleles in and out of a population due to the migration of individuals or gametes. While some populations are fairly stable, others experience more flux. Many plants, for example, send their pollen far and wide, by wind or by a pollinator, to pollinate other populations of the same species some distance away. Even a population that may initially appear to be stable, such as a pride of lions, experiences immigration (coming into) and emigration (departing from) as developing males leave their mothers to seek out a new pride with genetically unrelated females. The flow of individuals in and out of populations changes the population’s allele frequencies. When an individual migrates into a new population, it may sometimes carry alleles that were not present in that population before, introducing new genetic variation into that population. Alternatively, if an individual emigrates from (leaves) a population where it is the only individual with a particular allele, it can result in its original population losing genetic diversity. Migration between populations causes them to become more genetically similar to each other. For example, alleles that may be present in one population but not the other can become present in both populations when a migrating individual introduces that allele to the new population.
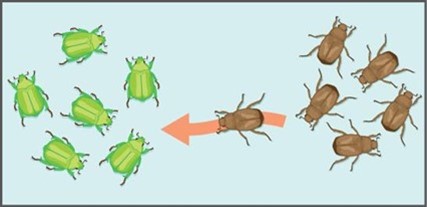
In either case, emigration or immigration, if it changes the allele frequency of a trait, it results in evolution. For example, if the trait in question is coloration and a population of beetles has only the variation or allele of green colour, the allele frequency will change if a brown beetle immigrates into this population. Evolution has occurred (Figure 1.3.5).
There are many examples of populations evolving through gene flow. One interesting case involves the global distribution of HIV resistance in humans. A mutation in the CCR5 gene provides resistance to certain forms of HIV, yet it is not the most common in regions with high rates of HIV and AIDS. This mutation is relatively recent—biochemical and geographic evidence suggests it originated in Northern Europe about 1,200 years ago. Interestingly, the mutation spread globally long before HIV became a threat to human health. Its distribution closely follows the migration patterns of the Vikings during the 9th to 11th centuries, suggesting they may have carried the mutation with them as they settled new territories. Why the mutation was common among Vikings is still uncertain—it may have offered a survival advantage, or it may have become widespread through the founder effect.
Natural Selection
We’ve already introduced natural selection, but let’s take a closer look.
Natural selection is the process by which individuals with traits better suited to their environment are more likely to survive and reproduce. Being better suited means they can more effectively overcome environmental challenges, known as selective pressures, such as predators, climate, food availability, or competition. Individuals with traits that help them cope with these pressures tend to leave more offspring, passing those beneficial traits to the next generation.
Over time, this process increases the frequency of advantageous traits in a population, leading to adaptation. An adaptation is a heritable trait that aids the survival and reproduction of an organism in its present environment. An adaptation is a “match” of the organism to the environment. Whether or not a trait is favourable depends on the environment at the time. The same traits do not always have the same relative benefit or disadvantage because environmental conditions can change. For example, finches with large bills benefited in one climate, while small bills were a disadvantage; in a different climate, the relationship reversed.

For natural selection to occur, there are three requirements:
- Variation. Multiple variants of genes must be present in a population for natural selection to produce any changes. Figure 1.3.6 shows a population of beetles that vary in their colour. In order for the population to evolve, the population must contain individuals with different colours. In the top panel, seven beetles are light-colored and 7 beetles are dark-colored.
- Heritability. For a trait to be subject to natural selection, it must be heritable. By heritable, we mean that the trait must be passed down from parents to their offspring through their DNA sequences. In the beetle example, light-colored beetles produce light offspring, and dark-colored beetles produce dark offspring.
- Differential reproductive success. An organism’s ability to produce a relatively high number of offspring is its reproductive success. In order to evolve via natural selection, an organism’s reproductive success must be directly affected by its traits. The number of offspring an individual produces in their lifetime is how scientists measure reproductive success. The more offspring, the higher the reproductive success. Some individuals will have no offspring, some a few, and some many. This is what we mean by differential reproductive success. In Figure 1.3.6, the beetles live on trees with dark-colored bark. Predators like birds can detect and kill light-colored beetles more easily than dark-colored beetles. This means that dark-colored beetles survive longer and therefore produce more offspring (have higher reproductive success than beetles with light coloration). We would say, “Dark-colored beetles are more fit”, where fitness is a measure of an individual’s relative reproductive success compared to others in the population.
There are several ways selection can affect population variation. The three general outcomes of natural selection include:
Stabilizing Selection
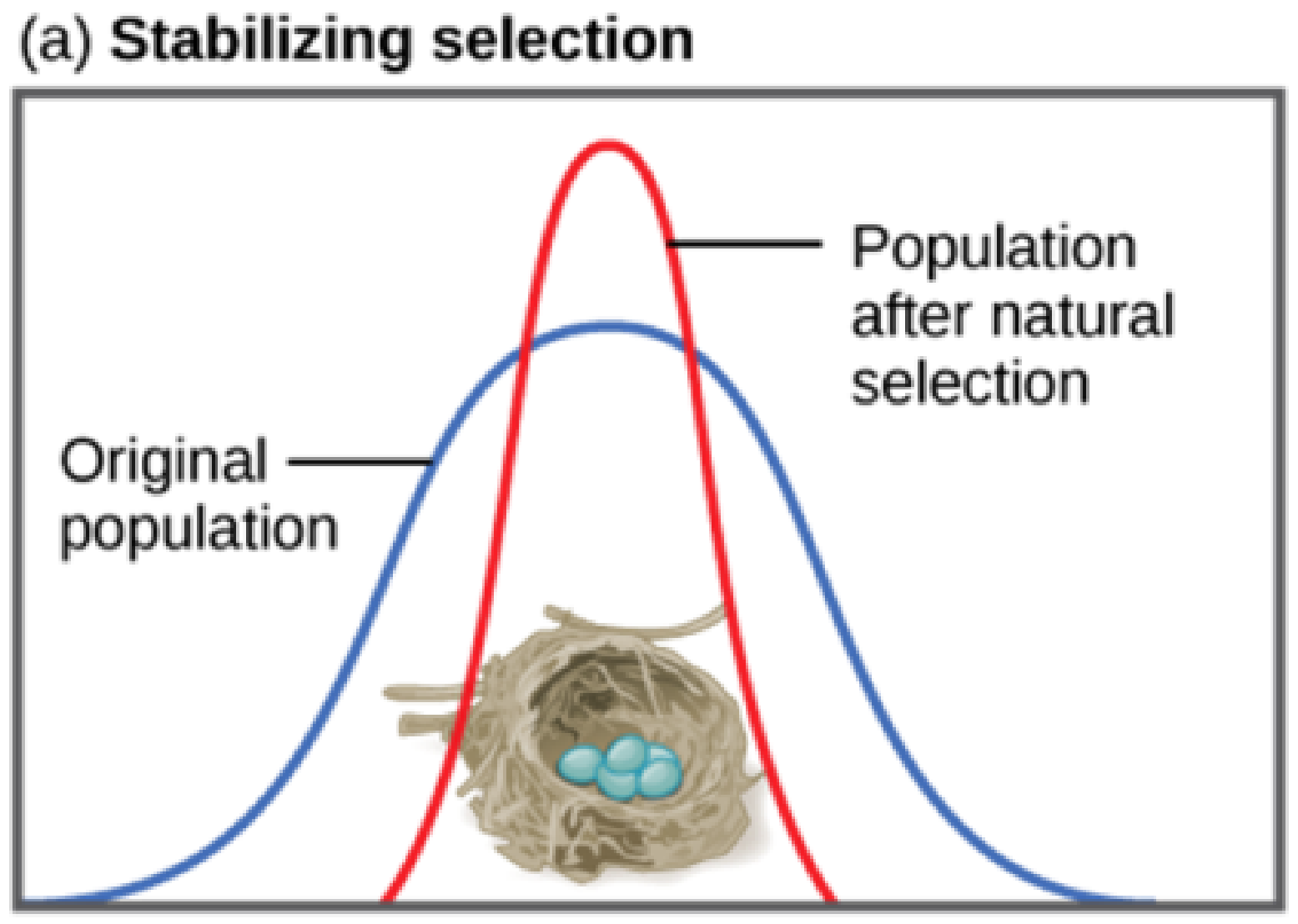
If natural selection favours an average phenotype, selecting against extreme variation, the population will undergo stabilizing selection. In a population of mice that live in the woods, for example, natural selection is likely to favour individuals that best blend in with the forest floor and are less likely to be spotted by predators. These camouflage mice are more adapted to that environment and will be more fit (reproducing more and leaving more descendants). Assuming the ground is a fairly consistent shade of brown, those mice whose fur is most closely matched to that colour will be most likely to survive and reproduce, passing on their genes for their brown coat. Mice that carry alleles that make them a bit lighter or a bit darker will stand out against the ground and be more likely to fall victim to predation. As a result of this selection, the population’s genetic variance will decrease.
Directional Selection
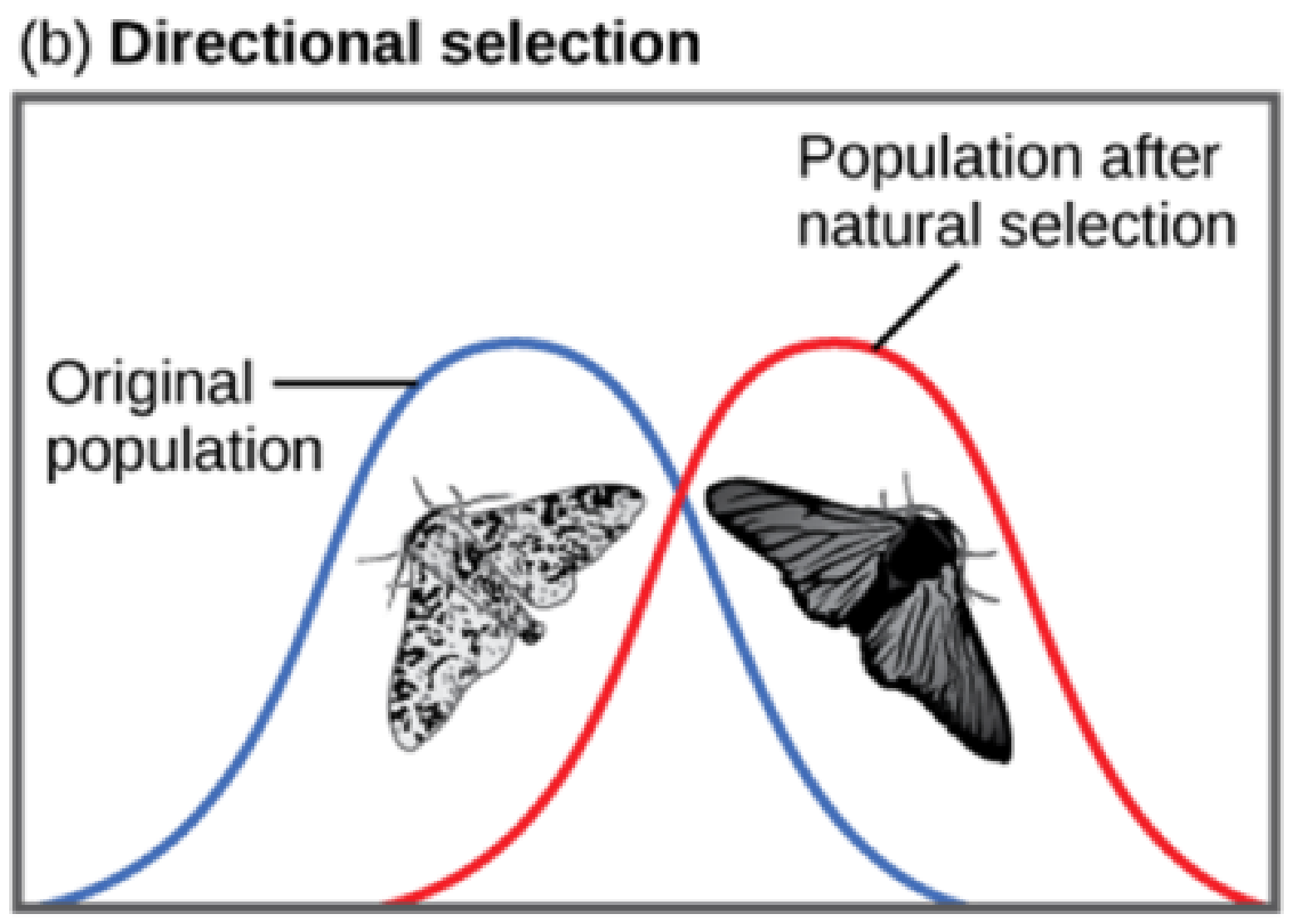
When the environment changes, populations will often undergo directional selection, which selects for phenotypes at one end of the spectrum of existing variation. A classic example of this type of selection is the evolution of the peppered moth in eighteenth- and nineteenth-century England. Prior to the Industrial Revolution, the moths were predominantly light in colour, which allowed them to blend in with the light-colored trees and lichens in their environment. But as soot began spewing from factories, the trees became darkened, and the light-colored moths became easier for predatory birds to spot. Over time, the frequency of the darker form of the moth increased because they had a higher survival rate in habitats affected by air pollution, as their darker coloration blended with the sooty trees. Similarly, the hypothetical mouse population may evolve to take on a different coloration if something were to cause the forest floor where they live to change colour. The result of this type of selection is a shift in the population’s genetic variance toward the new, fit phenotype.
Disruptive Selection
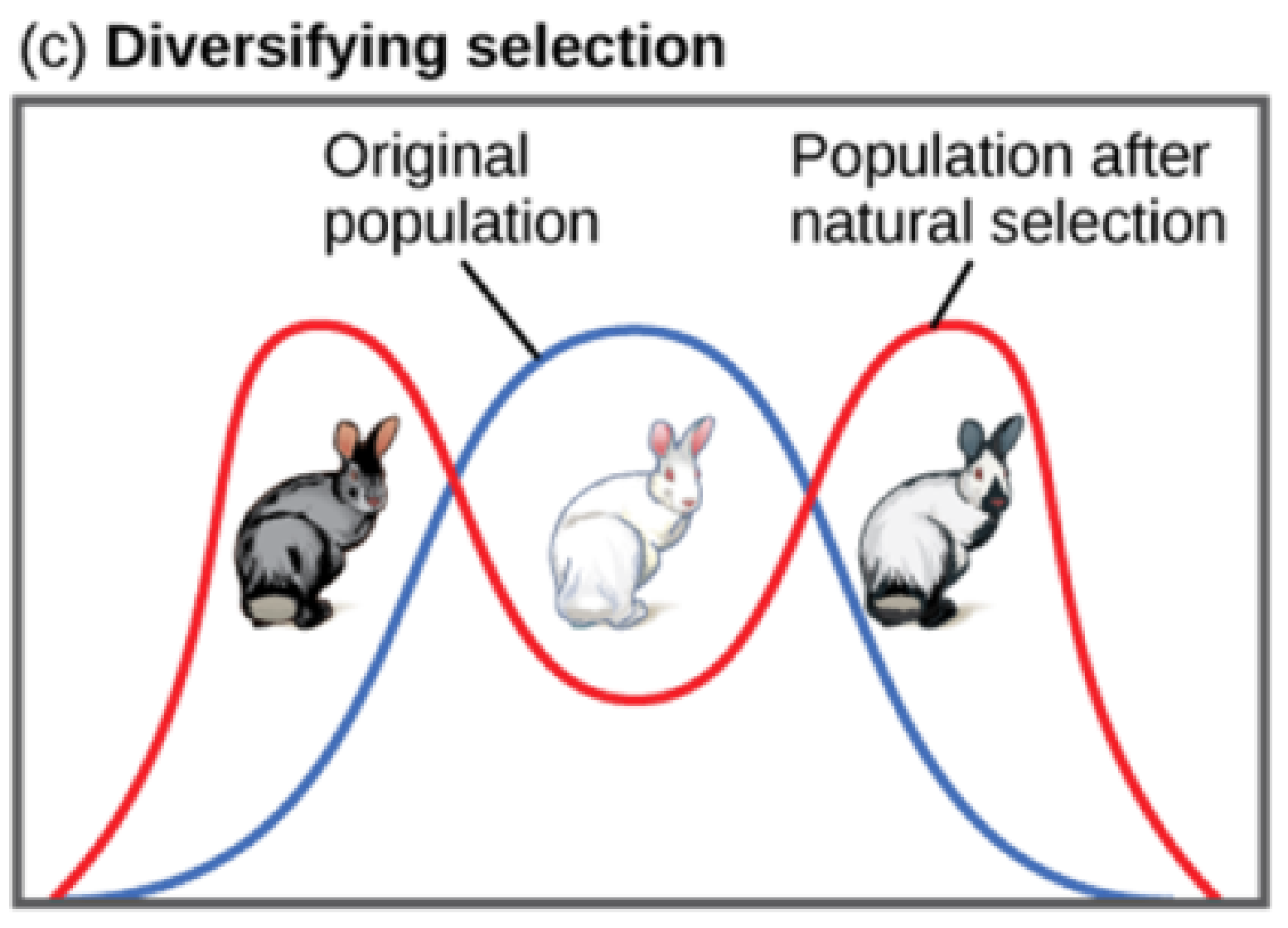
Sometimes two or more distinct phenotypes can each have their advantages and be selected for by natural selection, while the intermediate phenotypes are, on average, less fit. Known as disruptive (or diversifying) selection, this is seen in many populations of animals that have multiple male forms. Large, dominant alpha males obtain mates by brute force, while small males can sneak in for furtive copulations with the females in an alpha male’s territory. In this case, both the alpha males and the “sneaking” males will be selected for, but medium-sized males, which can’t overtake the alpha males and are too big to sneak copulations, are selected against. Diversifying selection can also occur when environmental changes favour individuals on either end of the phenotypic spectrum. Imagine a population of mice living at the beach where there is light-colored sand interspersed with patches of tall grass. In this scenario, light-colored mice that blend in with the sand would be favoured, as well as dark-colored mice that can hide in the grass. Medium-colored mice, on the other hand, would not blend in with either the grass or the sand and would thus be more likely to be eaten by predators. The result of this type of selection is increased genetic variance as the population becomes more diverse.
Frequency-Dependent Selection

Frequency-dependent selection is a type of natural selection that favours phenotypes that are either common (positive frequency-dependent selection) or rare (negative frequency-dependent selection). An interesting example of this type of selection is seen in a unique group of lizards of the Pacific Northwest. Male common side-blotched lizards come in three throat-colour patterns: orange, blue, and yellow. Each of these forms has a different reproductive strategy: orange males are the strongest and can fight other males for access to their females; blue males are medium-sized and form strong pair bonds with their mates; and yellow males (Figure 1.3.10) are the smallest, and look a bit like females, which allows them to sneak copulations. Like a game of rock-paper-scissors, orange beats blue, blue beats yellow, and yellow beats orange in the competition for females. That is, the big, strong orange males can fight off the blue males to mate with the blue’s pair-bonded females, the blue males are successful at guarding their mates against yellow sneaker males, and the yellow males can sneak copulations from the potential mates of the large, orange males.
In this scenario, orange males will be favoured by natural selection when the population is dominated by blue males, blue males will thrive when the population is mostly yellow males, and yellow males will be selected for when orange males are the most populous. As a result, populations of side-blotched lizards cycle in the distribution of these phenotypes—in one generation, orange might be predominant, and then yellow males will begin to rise in frequency. Once yellow males make up a majority of the population, blue males will be selected for. Finally, when blue males become common, orange males will once again be favoured.
Sexual Selection
Sexual selection is a form of natural selection that occurs when individuals with certain traits are more likely to attract mates and reproduce. These traits may not directly improve survival, but they increase reproductive success by making individuals more appealing to potential mates or more competitive in securing them.
In many species, males and females differ significantly in appearance beyond their reproductive organs—a phenomenon known as sexual dimorphism. Males are often larger and more elaborately adorned, like the peacock’s colourful tail, while females tend to be smaller and less decorated. This difference arises because, in many animal populations, a few males achieve most of the matings, while others may get none. Since eggs are energetically costly to produce and females often invest more in raising offspring, they tend to be more selective in choosing mates. This choosiness drives males to evolve traits that increase their chances of being selected, such as larger body size, brighter colours, or elaborate displays. As a result, sexual selection creates strong pressure on males to stand out and compete for limited mating opportunities.

Sexual dimorphism varies widely among species, of course, and some species are even sex-role reversed. In such cases, females tend to have a greater variance in their reproductive success than males and are correspondingly selected for the bigger body size and elaborate traits usually characteristic of males.
Knowledge Check
Text Description
- Genetic drift
- Artificial selection
- Natural selection
- Mutation
- Species can evolve over time
- Species were formed through natural selection
- Species were fixed and unchanging
- Species were created by natural forces
- The theory of natural selection
- The idea of creationism
- The belief that species could evolve over time
- The belief in extinct species
- Their beaks were identical on all islands
- They had wings adapted for long-distance flights
- Their beaks varied depending on their food sources
- They could only feed on seeds
- Thomas Malthus
- Charles Darwin
- Jean-Baptiste Lamarck
- Alfred Russel Wallace
- Heritability
- Variation
- Descent with modification
- Overproduction
- He worked with Lamarck on his evolutionary theory
- He independently developed a similar theory to Darwin’s
- He disproved the idea of species change
- He discovered the process of natural selection
- They had evolved unique adaptations to survive in the ocean
- They had no relationship to mainland species
- They were an example of extinction
- They were unchanged from their ancestors
Answers:
- c. Natural selection
- c. Species were fixed and unchanging
- c. The belief that species could evolve over time
- c. Their beaks varied depending on their food sources
- c. Jean-Baptiste Lamarck
- d. Overproduction
- b. He independently developed a similar theory to Darwin’s
- a. They had evolved unique adaptations to survive in the ocean
OpenAI. (2025). ChatGPT. [Large language model]. https://chat.openai.com/chat
Prompt: Create 8 multiple-choice questions using the following content
“Evolutionary Change Outside of Natural Selection” from Introductory Biology: Ecology, Evolution, and Biodiversity by Erica Kosal is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted. Modifications: Edited and reworded; Added additional images and knowledge check.
“11.2 Mechanisms of Evolution” from Biology and the Citizen by Colleen Jones is licensed under a Creative Commons Attribution 4.0 International License, except where otherwise noted. Modifications: Edited and reworded; Added additional images and knowledge check.
“2.10 Mechanisms of Evolution: Natural Selection” from Introduction to the Evolution & Biology of Sex by Katherine Furniss & Sarah Hammarlund is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, except where otherwise noted. Modifications: Edited and reworded; Added additional images.

{kind=link}
{kind=link}